HTTP 200 OK
Allow: GET, HEAD, OPTIONS
Content-Type: application/json
Vary: Accept
{
"count": 15,
"next": "https://plant-trends.bsmiab.org/articles/?format=api&page=2",
"previous": null,
"results": [
{
"id": 16,
"slug": "280-1733444100",
"featured": true,
"slider": false,
"issue": "Vol3 Issue1",
"type": "review_article",
"manuscript_id": "280-1733444100",
"recieved": "2024-12-06",
"revised": null,
"accepted": "2025-01-11",
"published": "2025-01-25",
"pdf_file": "https://plant-trends.bsmiab.org/media/pdf_file/2025/04/280-1733444100.pdf",
"title": "Harnessing plant–microorganism interactions for nano-bioremediation of heavy metals: Cutting-edge advances and mechanisms",
"abstract": "<p>Nano-bioremediation, an emerging eco-friendly strategy that integrates nanotechnology and biological processes to mitigate the contamination of heavy metals from the environment. To explores synergistic interactions between plants and microorganisms, focusing on their potential role in enhancing nano-bioremediation are highly demandable. This study focuses several key mechanisms including biosorption, bioaccumulation, biomineralization, and enzymatic reduction, the coordination of microorganisms and plants in tolerating and transforming toxic heavy metals into less toxic forms. The potential role of microorganism-assisted nanomaterials, including nano-biosorbents and nano-catalysts in phyto- and eco-environments were updated. This review also highlights recent studies on the significance of plant-microbe systems and nanomaterials in heavy metal remediation, challenges such as microbial survival, scalability, and ecological impacts were addressed, alongside potential solutions. Finally, this critical review provides new insights into harnessing plant–microorganism interactions for nano-bioremediation, presenting an eco-friendly approach to address global heavy metal pollution, and it shows a sustainable way of clean environment.</p>",
"journal_reference": "Plant Trends. 2025; 3(1): 01-12",
"academic_editor": "Md. Azizul Haque, PhD; Hajee Mohammad Danesh Science and Technology University, Bangladesh",
"cite_info": "Das K, Sarker A, et al. Harnessing plant–microorganism interactions for nano-bioremediation of heavy metals: Cutting-edge advances and mechanisms. Plant Trends. 2025; 3(1): 01-12.",
"keywords": [
"Pollution",
"Soil remediation",
"Environmental sustainability",
"Toxicity",
"Microorganisms"
],
"DOI": "10.5455/pt.2025.01",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>Bioremediation (BR) is one of the most efficient methods for removal of heavy-metal-from the contaminated soil and groundwater. This procedure is less harmful to the environment and more economical than conventional chemical and physical techniques, which are very costly and inefficient at low metal concentrations and result in large volumes of hazardous sludge [<a href=\"#r-1\">1</a>,<a href=\"#r-2\">2</a>]. The ability of microorganisms to breakdown contaminants is dependent on environmental conditions for growth and metabolism, which include favorable temperature, pH, and moisture [<a href=\"#r-3\">3</a>,<a href=\"#r-4\">4</a>]. Microorganisms are crucial for the removal of contaminants from soil, water, and sediments because of their benefits over alternative remediation techniques. These approaches of HMs removal are eco-friendly and cost effective. Additionally, they aid in the restoration of the natural ecosystem by preventing ongoing contamination [<a href=\"#r-5\">5</a>]. Also, the contamination of heavy metals (HMs) has become a severe hazard to the ecosystem [<a href=\"#r-6\">6</a>]. In the industrial sector, heavy metal is a profitable industry. Nevertheless, it is also a major environmental problem everywhere [<a href=\"#r-7\">7,8</a>]. Also, the environment contains natural, agricultural, solid waste, inland effluents, and air sources, for the additional heavy metal sources. Large portions of the earth have been polluted by mining, electroplating, metallurgical smelting, pesticide, and fertilizer use in agricultural fields [<a href=\"#r-9\">9</a>].</p>\r\n\r\n<p>Furthermore, HMs that find their way into the environment linger and seriously endanger creatures that come into proximity to them due to toxicity. Although very low quality is crucial for the biological operations of plants and animals, but in high dose is toxic and inhibit metabolism in other organisms [<a href=\"#r-10\">10</a>]. Toxic heavy metals, such as lead (Pb), cadmium (Cd), mercury (Hg), chromium (Cr), zinc (Zn), uranium (Ur), selenium (Se), silver (Ag), gold (Au), nickel (Ni), and arsenic (As), which are not useful to plants, inhibit plant growth, photosynthetic and enzymatic activities, and essential plant nutrition [<a href=\"#r-11\">11</a>]. Moreover, even at low concentrations, heavy metals are carcinogenic to humans [<a href=\"#r-12\">12</a>]. Conversely, bioremediators are biological agents used in bioremediation to help to clean contaminated sites. Among the most often used bioremediators are bacteria, archea, and fungi [<a href=\"#r-13\">13</a>]. The application of bioremediation, a biotechnological method that uses microorganisms to solve and eliminate environmental concerns caused by different pollutants through biodegradation.</p>\r\n\r\n<p>Scientists are coming to an agreement on how to reduce pollutant release and mitigate their impacts using living creatures such as plants, which is known as phytoremediation or bacteria, which is referred to as bioremediation [<a href=\"#r-14\">14</a>]. To address these issues, biological methods such as biosorption, bioaccumulation, biodegradation, and bioremediation are used to remove heavy metal ions, providing an appealing alternative to physicochemical methods [<a href=\"#r-15\">15</a>]. These methods are potentially simple, low cost, more effective, ecofriendly, and a self-sustaining option for wastewater amelioration, which is gaining new attention nowadays [<a href=\"#r-16\">16</a>]. Also, the chemical methods for heavy metal remediation pose health hazards such as toxicity and environmental risks, coupled with limitations like high costs and inefficiency under varying conditions. In contrast, microorganism-assisted approaches offer eco-friendly, sustainable, and cost-effective alternatives, leveraging natural mechanisms for safe and effective HMs removal [<a href=\"#r-2\">2</a>,<a href=\"#r-17\">17</a>].</p>\r\n\r\n<p>The goal of this review is to explore current trends in the application/role of microorganisms in bioremediation with their interactions for nano-bioremediation, find the necessary background information to fill in any gaps in this theme area. In this study, the nono-bioremediation strategy provides a sustainable way for removing or minimizing HMs from the plants, soils and environments.</p>"
},
{
"section_number": 2,
"section_title": "SOURCES OF HEAVY METALS",
"body": "<p>Natural resources, such as air, soil, and water ecosystems, have been discovered to contain HMs, agricultural chemicals, industrial solvents (especially chlorinated solvents), and other types of pollution [<a href=\"#r-18\">18</a>]. The use of HMs by industry and agricultural sectors has resulted in massive amounts of HMs being relinquished and disposed of inadvertently in most ecosystems [<a href=\"#r-19\">19</a>]. Several agricultural practices, such as the irreversible use of urban sewage sludge, industry-based practices, such as composting and burning of garbage in a variety of techniques, and vehicle emissions unintentionally introduce toxic metals (Cd, Cr, Pb, Hg, As, Cu, Zn, and Ni) into soils (<a href=\"#figure1\">Figure 1</a>). Motor vehicle emissions (Pb) [<a href=\"#r-20\">20</a>], engine wear (Cd, Cu, and Ni) [<a href=\"#r-21\">21</a>], and tire abrasion (e.g., Zn). According to Hunter [<a href=\"#r-22\">22</a>], arsenic could have positive effects on gene silencing and methionine metabolism in animals. Arsenic and cadmium are naturally present in very small amounts in the Earth's crust and probably weren't conscripted during evolutionary processes because they are less abundant than P and Zn, which occupy adjacent columns in the periodic table, respectively [<a href=\"#r-23\">23</a>].</p>\r\n\r\n<p>The accumulation of potentially harmful quantities of As and Cd in soils is largely due to anthropogenic activity [<a href=\"#r-24\">24</a>]. Metal toxicity is critical for living organisms, including microbes, plants, animals, and humans. However, the toxicity varies depending on the organism, while most of the 80 metals detected are essential for human functioning biology (e.g., Fe, Mg, Zn), others, such as Pb, Hg, and Cd, are among the oldest human toxicants [<a href=\"#r-25\">25</a>]. A few heavy metals, including Fe, Cu, and Zn, are necessary microelements, whereas others, like Cd and Pb, have no beneficial function and are harmful even at low quantities (<a href=\"#figure1\">Figure 1</a>). The contamination of soil and aquatic ecosystems is important, because metals are not biodegradable like other organic contaminants, they accumulate in terrestrial, aquatic, and marine ecosystems [<a href=\"#r-26\">26</a>].</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"507\" src=\"/media/article_images/2025/46/02/280-1733444100-Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1.</strong> Sources of HMs in the environment. The figure illustrates the various anthropogenic and natural sources of heavy metals, such as industrial discharge, agricultural runoff, natural mineral deposits, soil waste and toxic chemicals.</figcaption>\r\n</figure>\r\n</div>"
},
{
"section_number": 3,
"section_title": "HEAVY METALS CONTAMINATION AND PUBLIC HEALTH RISK",
"body": "<p>HMs contamination problem is one of the major concerns worldwide. HMs toxicity varies due to their toxicity levels. The HMs are fluently added to environments from different sources (<a href=\"#figure1\">Figure 1</a>), and those can easily deposit into living organisms [<a href=\"#r-27\">27</a>]. They are all metals that have an atomic weight higher than that of iron (55.8 gmol<sup>–1</sup>), and they are found in the environment. However, some metals have an atomic weight lower than that of Fe. For example, Cr, and some metalloids, like As and Se, are also called "heavy metals" [<a href=\"#r-28\">28</a>]. Many types of HMs can be micronutrients for humans. These include Cu, Fe, Mn, Mo, Zn, and Ni. They can also be toxic to humans if being exposed to them for a longer period, like Hg, Pb, Cd, Cu, Ni, and Co. Contamination by HMs has a lot of bad effects, not just on animals and plants but also on human health [<a href=\"#r-29\">29</a>]. As an example, Zn is a component of a few enzymes, such as enzymes that break down carbohydrates, proteins, and peptides, as well as enzymes that make RNA and ribosomes in plants. Copper helps plants do a lot of things, like photosynthesis, respiration, carbohydrate distribution, nitrogen and cell wall metabolism, seed production, and disease resistance, but at high concentrations, these metals can harm cells [<a href=\"#r-30\">30</a>]. The Cd is highly toxic for biological processes and very harmful to organisms when it builds up even in low quantities. Deposition of HMs depends on metal ions specific ion-binding processes to specific locations, and cellular structure<strong> </strong>variability. HMs ions, have a strong electrostatic attraction and great binding affinities with these same locations. The toxicity outputs of HMs are critical for basic genetic molecules of organisms. The structures and biomolecules, such as cell wall enzymes, DNA, and RNA, become unstable because of this, which is ultimately responsible for occurring mutations at molecular levels, as a result altering genetic levels, physiological difficulties, illnesses, and even cancer [<a href=\"#r-31\">31</a>].</p>"
},
{
"section_number": 4,
"section_title": "HEAVY METAL TOXICITY IN PLANTS, SOIL, AND ENVIRONMENT",
"body": "<p>Globally, due to the persistence, high toxicity, and recalcitrant nature, metal contamination has now become a serious concern in plants, soil, and the environment (<a href=\"#figure2\">Figure 2</a>). These toxic metals have posed a serious threat to the environmental stability and health of all living organisms [<a href=\"#r-2\">2</a>]. Like other living organisms, plants are not resistant to high concentrations of HMs in the air due to human activities and environment. Trace amounts of HMs enhance plants by acting as essential micronutrients. According to Lopez-Vargas et al. [<a href=\"#r-32\">32</a>], copper can improve the flavor and color of floral arrangements, fruits, and vegetables by increasing sugar content in plants. Zn is an essential component of the enzymatic system as well as the metabolic processes of plants [<a href=\"#r-33\">33</a>], and photosynthetic compounds are particularly influenced by HMs [<a href=\"#r-34\">34</a>]. Heavy metals have been shown to accumulate in plants, where they interfere with the normal metabolic and biological processes in the plant, and eventually leading to severe yield losses [<a href=\"#r-35\">35</a>].</p>\r\n\r\n<p>HMs prevent seed germination by adversely influencing the processes, which in turn reduces the establishment of the entire stand [<a href=\"#r-36\">36</a>]. Due to the high concentration of malondialdehyde (MDA) and H<sub>2</sub>O<sub>2</sub>, HM also impairs the water status of plants, damages the stability of their membranes, and increases the loss of crucial osmolytes. It shows that different chemical, physical, and biological methods for HMs removal from soils have been in practice globally. Plants consume and accumulate HM that is present in the soil at very high concentrations and eventually reaches human nutrition through the food chain [<a href=\"#r-37\">37</a>]. Recently, microbes have gained a lot of attention from scientists worldwide. Biosorption, bioaccumulation, biovolatilization, biomineralization, oxidation and reduction, bioleaching, and the synthesis of bio-surfactants are some of the methods by which the bacteria extract the heavy metals from the soil [<a href=\"#r-17\">17</a>].</p>\r\n\r\n<p>Plants and microorganisms are used as biological methods to treat HMs containing polluted soils [<a href=\"#r-38\">38</a>]. Hints, there are also certain drawbacks to these techniques in terms of long durations, environmental sensitivity, and toxicity of contaminants [<a href=\"#r-39\">39</a>]. High levels of HMs in soil and water are representative examples of human activities which have a significant effect on the environment and present a huge risk [<a href=\"#r-40\">40,41</a>]. According to Abd Elnabi et al. [<a href=\"#r-42\">42</a>], living plants and animals are at serious risk due to the toxic HM's persistence in the soil environment. For terrestrial plants, the primary points of contact with hazardous heavy metals (HMs) are the roots [<a href=\"#r-43\">43</a>]. Additionally, microbes lower the concentration of heavy metals (HMs) in soil. For example, <em>Aspergillus niger</em> shown a notable capacity to bioaccumulate Cd and Cr [<a href=\"#r-44\">44</a>], and <em>Stenotrophomonas rhizophila</em> also considerably reduced Pb and Cu by 76.9% and 83.4%, respectively [<a href=\"#r-45\">45</a>]. Microbes have a large surface area to adsorb the HMs because of their small size, which lowers the total amount of HMs available [<a href=\"#r-46\">46</a>]. Soil biology is indispensable with respect to soil quality maintenance, which again is very important for sustainable agriculture. Human activities have emerged as the prime source of HMs and have disturbed the soil microbes, soil fertility, and productivity [<a href=\"#r-47\">47</a>].</p>\r\n\r\n<p>However, their bioaccumulation and biomagnification attributes in the food chain are highly threatening to the environment due to their accumulation in soil and plants [<a href=\"#r-48\">48</a>]. Application of chemical fertilizers and pesticides can enhance the risk of HMs contamination in the soil. The chemical toxicity outputs lead to build-up in crop tissue grown in the contaminated soil [<a href=\"#r-49\">49</a>]. Microbial bioremediation methods have recently demonstrated significant promise in cleaning up contaminated soils. One type of green technology is the utilization of microorganisms' metabolic processes to remove heavy metal contamination, and most HMs have been classified as hazardous overall.</p>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"310\" src=\"/media/article_images/2025/46/02/280-1733444100-Figure2.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 2.</strong> Toxicity of HMs in plants, soil, and environment. The figure represents the toxic effects of HMs on plant physiology, soil fertility, and overall environmental health. The figure highlights specific mechanisms of toxicity such as disruption of enzymatic activities and oxidative stress in plants, alongside soil degradation.</figcaption>\r\n</figure>\r\n</div>"
},
{
"section_number": 5,
"section_title": "NANO-BIOREMEDIATION OF HEAVY METALS USING MICROORGANISMS",
"body": "<p>Nanoparticles-based bioremediation is an emerging and highly efficient approach for large-scale environmental cleanup, minimizing toxic repercussions. In the relentless march of technology, bioremediation has evolved into nano-bioremediation, employing nanoparticles and microbes to offer eco-friendly solutions for tackling hazardous environmental pollutants (<a href=\"#figure3\">Figure 3</a>).</p>\r\n\r\n<p>Microorganisms-based HMs removing or minimizing are more efficient approaches compared to traditional methods [<a href=\"#r-50\">50</a>]. Metals like As, Cd, and Pb are highly toxic even at low concentrations [<a href=\"#r-51\">51</a>]. Microbial bioremediation immobilizes these metals; for example, <em>Morganella psychrotolerans</em> produces silver nanoparticles for heavy metal removal [<a href=\"#r-52\">52</a>], while iron oxide nanoparticles with polyvinyl pyrrolidone (PVP) and <em>Halomonas</em> sp. effectively remediate Pb and Cd [<a href=\"#r-53\">53</a>]. Additionally, silica nanoparticles, <em>Pseudomonas aeruginosa</em>, and graphene oxide remove polycyclic aromatic hydrocarbons (PAHs) [<a href=\"#r-54\">54</a>], and <em>Halomonas</em> immobilized with magnetic NPs degraded Pd metal [<a href=\"#r-55\">55</a>]. Moreover, Bacteria synthesize diverse nanoparticles used for immobilization and mobilization of metals [<a href=\"#r-56\">56</a>], with strains like <em>Bacillus cereus</em> (PMBL-3) and <em>Lactobacillus macroides</em> (PMBL-7) effectively remediating HMs such as Cd, Cr, Pd, and Cu [<a href=\"#r-57\">57</a>]. Interestingly, Myconanotechnology employs fungi for bioremediation (<a href=\"#Table-1\">Table 1</a>), with various mushroom species effectively remediating soil contaminants [<a href=\"#r-58\">58</a>, <a href=\"#r-59\">59</a>]. Fungi such as <em>Fusarium solani</em> are increasingly being used in the process of nanoparticle manufacturing, because of their resistance to heavy metals [<a href=\"#r-60\">60</a>], while <em>Trichoderma harzianum</em> degrades pentachlorophenol, and <em>Cryptococcus</em> sp. displays resilience to HMs [<a href=\"#r-61\">61</a>].</p>\r\n\r\n<p>The chemical structure and molecular weight of microplastics (MPs), along with environmental conditions, influence microbial-driven degradation. This process involves biodeterioration, bio-fragmentation, biosynthesis, and mineralization [<a href=\"#r-82\">82</a>]. Although the precise mechanism of <em>Pseudomonas</em> sp. in degrading MPs particles remains elusive, research suggests the involvement of chitinase degradation [<a href=\"#r-83\">83</a>]. Microorganisms utilize metal-based nanoparticles (MNPs) as a carbon source for growth, aiding in the degradation of high molecular-weight plastics [<a href=\"#r-84\">84</a>]. The microbial degradation of plastic fragments offers a green solution, but controlling changes in plastic pollution relies on various factors. Therefore, the utilization of effective microbes can be a suitable approach for eliminating MNPs [<a href=\"#r-81\">81</a>, <a href=\"#r-85\">85</a>]. However, microbial degradation of MNPs is still in its infancy, as the characterization process is very slow, and incomplete mineralization is another limitation [<a href=\"#r-86\">86</a>]. MNPs present in municipal solid waste (MSW) can harbor various microinorganic and organic pollutants, posing risks to the environment and human health as they enter the food chain [<a href=\"#r-87\">87</a>]. Indigenous microbial communities within MSW and sewage sludge exhibit plastic degradation capabilities. Recent studies have shown that mesophilic <em>Stenotrophomonas panacihumi</em> can convert polypropylene (PP) into low and high-molecular-weight forms after 90 days [<a href=\"#r-88\">88</a>]. The persistence of antibacterial nanoparticles beyond a threshold poses a threat to soil microbes, potentially inhibiting nitrogen-fixing microbes, leading to stunted plant growth and reduced production [<a href=\"#r-89\">89</a>].<br />\r\nIn the perspective of environmental bioremediation, leveraging nanotechnology alongside microbial assistance emerges as a highly efficient strategy, as evidenced by the effective removal of HMs and pollutants from soil and wastewater. While nano-bioremediation offers promising solutions, the persistence of antibacterial nanoparticles poses challenges to soil microbial communities and ecosystems. Despite advancements, microbial degradation of micro-nano plastics remains in its nascent stages, highlighting the need for further research to optimize biodegradation processes and minimize environmental risks.</p>\r\n\r\n<div id=\"figure3\">\r\n<figure class=\"image\"><img alt=\"\" height=\"166\" src=\"/media/article_images/2025/46/02/280-1733444100-Figure3.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 3.</strong> Role of nanoparticles in environmental clean-up. The figure depicts the sources of nanoparticles and their mechanisms of action in remediation of contaminated soil, water, and air/environment. It outlines the enzymatic processes involved and the effectiveness of various nanoparticles in sequestering or degrading heavy metal pollutants. These extended captions provide a comprehensive description of each figure, ensuring that readers can fully understand the implications and contexts of the visual data presented.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p style=\"text-align:center\"><strong>Table 1.</strong> Potential implication of microorganisms for bioremediation of HMs from plants, soils and environments.</p>\r\n\r\n<div id=\"Table-1\">\r\n<p style=\"text-align:center\"><a href=\"https://plant-trends.bsmiab.org/table/280-1733444100-table1/\">Table-1</a></p>\r\n</div>"
},
{
"section_number": 6,
"section_title": "LIMITATIONS AND FUTURE PROSPECTS",
"body": "<p>The application of microbes in remediation of HMs is known as nano-bioremediation and has drawn significant attention due to its efficient and environmentally friendly nature. However, several limitations have also been noticed during its widespread application. Saharan et al. [<a href=\"#r-90\">90</a>] observed, the HMs toxicity in microbial communities affect the metabolic activity of these microorganisms and consequently their overall efficacy. In contrast, the environmental persistence of engineered NPs expands concerns about their potential ecotoxicological impacts, including bioaccumulation and non-target effects [<a href=\"#r-91\">91</a>]. Laboratory findings often struggle to translate to field conditions due to variations in soil composition, pH, temperature, and competing ions, alongside unpredictable interactions among nanoparticles, plants, and microbes that are not fully understood [<a href=\"#r-92\">92</a>, <a href=\"#r-93\">93</a>]. The characteristics of engineered nanoparticles can change due to agglomeration or dissolution, affecting their reactivity and potential toxicity [<a href=\"#r-94\">94</a>]. Additionally, the long-term impacts of these nanoparticles, such as their accumulation in plant tissues or migration into water sources, remain poorly characterized. The scalability of nanoparticles is limited by expensive synthesis techniques and the high costs of large-scale field applications, especially in resource-constrained regions. Nanoparticles can introduce new environmental contaminants and potentially disrupt soil microbiomes, altering nutrient cycles and ecosystem balance due to high concentrations. The adoption of nanotechnology is complicated by inconsistent regulations across different regions and the absence of standardized protocols for assessing environmental safety and long-term impacts [<a href=\"#r-95\">95</a>].</p>\r\n\r\n<p>The environmentally friendly biodegradable nanoparticles will alleviate the worries about persistence and eco-toxicity effects [<a href=\"#r-68\">68</a>]. Integration with omics technologies, particularly genomics and proteomics, will provide deeper insight into the interaction of microbes with nanoparticles, thereby facilitating the design of tailored bioremediation strategies for specific contaminants [<a href=\"#r-96\">96</a>]. Real-time monitoring systems are integrated with nano-bioremediation, also including biosensors that enhance process efficiency while ensuring site-specific applicability [<a href=\"#r-17\">17</a>]. Advancing the field of nano-bioremediation of heavy metals through plant-microorganism interactions involves several promising directions. Elucidating mechanisms at the molecular level using comprehensive omics approaches and real-time monitoring techniques, such as synchrotron-based spectroscopy, can support deep insights into processes involved in HM and nanoparticle transformations.</p>\r\n\r\n<p>Developing eco-friendly and cost-effective nanomaterials through green synthesis methods and creating biodegradable nanoparticles can minimize environmental impact and production costs. Integrating nano-bioremediation with different remediation strategies, such as phytoremediation and chemical techniques, can enhance contaminant removal efficiency and manage complex pollution scenarios more effectively. Conducting field trials and validating long-term effectiveness under various conditions is crucial for assessing the feasibility and sustainability of nano-bioremediation methods. Additionally, the application of advanced models and tools, including AI and machine learning, can optimize remediation processes and customize approaches based on local environmental conditions. Harmonization and the establishment of standardized risk-benefit analyses are essential for gaining broader acceptance and ensuring the responsible implementation of nano-bioremediation technologies. These efforts will help refine nano-bioremediation strategies, making them more efficient, scalable, and environmentally safe. Therefore, the application of advanced nano-based tools and sustainable restoration of ecosystems from HMs-contaminated environments would be supportive for making smart and HMs-free green environments.</p>"
},
{
"section_number": 7,
"section_title": "CONCLUSIONS",
"body": "<p>This study explores effective strategies for microorganism-assisted HMs remediation, the role of plant-microbe interactions for nano-bioremediation of HMs, and strategies for transforming toxic HMs into less toxic forms. This study updates the techniques like biomineralization, biostimulation, and mycoremediation, discusses how microbe-assisted phytoremediation is crucial for minimizing HMs contamination, and shows sustainable solutions for maintaining of HMs toxicity. This review also highlights the prospect of using nano-environmental biotechnology tools for mitigating HMs toxicity in plants, soils, and environments. This updated study with eco-friendly approaches to HMs removal from environments could be useful for minimizing HMs toxicity and converting to smart-green earth.<strong> </strong></p>"
},
{
"section_number": 8,
"section_title": "ACKNOWLEDGEMENTS",
"body": "<p>The authors would like to acknowledge the College of Agriculture, Food and Environmental Sciences, California Polytechnic State University, San Luis Obispo, USA for providing research facilities and assistance.</p>"
},
{
"section_number": 9,
"section_title": "AUTHOR CONTRIBUTIONS",
"body": "<p>KD conceived the research plan, and supervised the study. KD, MAAM, and AS wrote the initial draft of the manuscript. KD, FMA, and SD edited the manuscript. All authors have approved the final version.</p>"
},
{
"section_number": 10,
"section_title": "CONFLICTS OF INTEREST",
"body": "<p>There is no conflict of interest among the authors.</p>"
}
],
"figures": [
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2025/46/02/280-1733444100-Figure1.jpg",
"caption": "Figure 1. Sources of HMs in the environment. The figure illustrates the various anthropogenic and natural sources of heavy metals, such as industrial discharge, agricultural runoff, natural mineral deposits, soil waste and toxic chemicals.",
"featured": false
},
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2025/46/02/280-1733444100-Figure2.jpg",
"caption": "Figure 2. Toxicity of HMs in plants, soil, and environment. The figure represents the toxic effects of HMs on plant physiology, soil fertility, and overall environmental health. The figure highlights specific mechanisms of toxicity such as disruption of enzymatic activities and oxidative stress in plants, alongside soil degradation.",
"featured": true
},
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2025/46/02/280-1733444100-Figure3.jpg",
"caption": "Figure 3. Role of nanoparticles in environmental clean-up. The figure depicts the sources of nanoparticles and their mechanisms of action in remediation of contaminated soil, water, and air/environment. It outlines the enzymatic processes involved and the effectiveness of various nanoparticles in sequestering or degrading heavy metal pollutants. These extended captions provide a comprehensive description of each figure, ensuring that readers can fully understand the implications and contexts of the visual data presented.",
"featured": false
}
],
"authors": [
{
"id": 75,
"affiliation": [
{
"affiliation": "College of Agriculture, Food and Environmental Sciences, California Polytechnic State University, CA 93407, USA"
}
],
"first_name": "Kallol",
"family_name": "Das",
"email": "kdas01@calpoly.edu",
"author_order": 1,
"ORCID": "https://orcid.org/0000-0003-0906-3983",
"co_first_author": false,
"co_author": false,
"corresponding": true,
"corresponding_author_info": "Kallol Das, PhD; College of Agriculture, Food and Environmental Sciences, California Polytechnic State University, CA- 93407, USA. \r\nEmail: kdas01@calpoly.edu",
"article": 16
},
{
"id": 76,
"affiliation": [
{
"affiliation": "National Institute of Agricultural Sciences, Rural Development Administration, Jeollabuk-do 55365, Republic of Korea"
}
],
"first_name": "Aniruddha",
"family_name": "Sarker",
"email": null,
"author_order": 2,
"ORCID": "https://orcid.org/0000-0001-6751-7301",
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 16
},
{
"id": 77,
"affiliation": [
{
"affiliation": "Civil, Construction, and Environmental Engineering, The University of Alabama, Tuscaloosa, AL 35487, USA"
}
],
"first_name": "Md Abdullah Al",
"family_name": "Masud",
"email": null,
"author_order": 3,
"ORCID": "https://orcid.org/0000-0002-8923-6137",
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 16
},
{
"id": 78,
"affiliation": [
{
"affiliation": "College of Agriculture, Food and Environmental Sciences, California Polytechnic State University, CA 93407, USA"
}
],
"first_name": "Shunping",
"family_name": "Ding",
"email": null,
"author_order": 4,
"ORCID": "https://orcid.org/0000-0001-6930-2806",
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 16
},
{
"id": 79,
"affiliation": [
{
"affiliation": "Department of Plant Pathology, Faculty of Agriculture, Sher-e-Bangla Agricultural University, Sher-e-Bangla Nagar, Dhaka, Bangladesh"
}
],
"first_name": "F. M.",
"family_name": "Aminuzzaman",
"email": null,
"author_order": 5,
"ORCID": "https://orcid.org/0000-0003-4804-0100",
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 16
}
],
"views": 561,
"downloads": 196,
"references": [
{
"id": 564,
"serial_number": 1,
"reference": "Ayangbenro AS, Babalola OO. A new strategy for heavy metal polluted environments: a review of microbial biosorbents. International Journal of Environmental Research and Public Health. 2017;14:94.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 565,
"serial_number": 2,
"reference": "Tang H, Xiang G, et al. Microbial mediated remediation of heavy metals toxicity: mechanisms and future prospects. Frontiers in Plant Science. 2024;15:1420408.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 566,
"serial_number": 3,
"reference": "Angon PB, Islam MS, et al. Sources, effects and present perspectives of heavy metals contamination: soil, plants and human food chain. Heliyon. 2024;10:e28357.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 567,
"serial_number": 4,
"reference": "Verma JP, Jaiswal DK. Book review: Advances in biodegradation and bioremediation of industrial waste. Frontiers in Microbiology. 2016;6:1555.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 568,
"serial_number": 5,
"reference": "Demnerova K, Mackova M, et al. Two approaches to biological decontamination of groundwater and soil polluted by aromatics characterization of microbial populations. International Microbiology. 2005;8:205–11.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 569,
"serial_number": 6,
"reference": "Deepa CN, Suresha S. Biosorption of lead (II) from aqueous solution and industrial effluent by using leaves of Araucaria cookii: application of response surface methodology. Journal of Environmental Science, Toxicology and Food Technology. 2014;8(7):67–79.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 570,
"serial_number": 7,
"reference": "Siddiquee S, Rovina K, et al. Heavy metal contaminants removal from wastewater using the potential filamentous fungi biomass: a review. Journal of Microbial & Biochemical Technology. 2015;7(6):384–95.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 571,
"serial_number": 8,
"reference": "Igiri BE, Okoduwa SI, et al. Toxicity and bioremediation of heavy metals contaminated ecosystem from tannery wastewater: a review. Journal of Toxicology. 2018;1–16.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 572,
"serial_number": 9,
"reference": "Zhang WJ, Jiang FB, et al. Global pesticide consumption and pollution: with China as a focus. PIAEES. 2011;1(2):125–44.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 573,
"serial_number": 10,
"reference": "Ojuederie OB, Babalola OO. Microbial and plant-assisted bioremediation of heavy metal polluted environments: a review. International Journal of Environmental Research and Public Health. 2017;14:1504.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 574,
"serial_number": 11,
"reference": "Nematian MA, Kazemeini F. Accumulation of Pb, Zn, C, and Fe in plants and hyperaccumulator choice in Galali iron mine area, Iran. International Journal of Agriculture and Crop Sciences. 2013;5:426.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 575,
"serial_number": 12,
"reference": "Dixit R, Malaviya D, et al. Bioremediation of heavy metals from soil and aquatic environment: an overview of principles and criteria of fundamental processes. Sustainability. 2015;7:2189–212.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 576,
"serial_number": 13,
"reference": "Strong PJ, Burgess JE. Treatment methods for wine-related and distillery wastewaters: a review. Bioremediation Journal. 2008;12:70–87.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 577,
"serial_number": 14,
"reference": "Conesa HM, Evangelou MWH, et al. A critical view of current state of phytotechnologies to remediate soils: still a promising tool?. The Scientific World Journal. 2012;2012:173829.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 578,
"serial_number": 15,
"reference": "Wang J, Chen C. Biosorbents for heavy metal removal and their future. Biotechnology Advances. 2009;27:195–226.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 579,
"serial_number": 16,
"reference": "Jothimani P, Bhaskaran A. Assessing the azoreductase enzyme activity of bacterial cultures used for decolourisation. Journal of Ecotoxicology & Environmental Monitoring. 2003;13(3):179–83.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 580,
"serial_number": 17,
"reference": "Ur Rahman S, Qin A, et al. Pb uptake, accumulation, and translocation in plants: plant physiological, biochemical, and molecular response: a review. Heliyon. 2024;10:e27724.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 581,
"serial_number": 18,
"reference": "Rajkumar M, Ae N, et al. Potential of siderophore-producing bacteria for improving heavy metal phytoextraction. Trends in Biotechnology. 2010;28:142–9.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 582,
"serial_number": 19,
"reference": "Ansari MI, Malik A. Biosorption of nickel and cadmium by metal-resistant bacterial isolates from agricultural soil irrigated with industrial wastewater. Bioresource Technology. 2007;98:3149–53.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 583,
"serial_number": 20,
"reference": "Sutherland RA, Day JP, et al. Lead concentrations, isotope ratios, and source apportionment in road-deposited sediments, Honolulu, Oahu, Hawaii. Water, Air, and Soil Pollution. 2003;142:165–86.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 584,
"serial_number": 21,
"reference": "Ozkutlu F, Turan M, et al. Assessment of heavy metal accumulation in the soils and hazelnut (Corylus avellana L.) from Black Sea coastal region of Turkey. Asian Journal of Chemistry. 2009;21:4371–88.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 585,
"serial_number": 22,
"reference": "Hunter P. A toxic brew we cannot live without. EMBO Reports. 2008;9(1):15–8.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 586,
"serial_number": 23,
"reference": "Zhao FJ, Ma JF, et al. Arsenic uptake and metabolism in plants. New Phytologist. 2009;181(4):777–94.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 587,
"serial_number": 24,
"reference": "Verbruggen N, Hermans C, et al. Mechanisms to cope with arsenic or cadmium excess in plants. Current Opinion in Plant Biology. 2009;12:1–9.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 588,
"serial_number": 25,
"reference": "Igwe JC, Nnorom IC, et al. Kinetics of radionuclides and heavy metals behavior in soils: implications for plant growth. African Journal of Biotechnology. 2005;4(B):1541–7.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 589,
"serial_number": 26,
"reference": "Ortega-Larrocea MP, Siebe C, et al. Mycorrhizal inoculum potential of arbuscular mycorrhizal fungi in soils irrigated with wastewater for various lengths of time, as affected by heavy metals and available P. Applied Soil Ecology. 2007;37:129–38.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 590,
"serial_number": 27,
"reference": "Zhuang W, Gao X. Acid-volatile sulfide and simultaneously extracted metals in surface sediments of the southwestern coastal Laizhou Bay, Bohai Sea: concentrations, spatial distributions and the indication of heavy metal pollution status. Marine Pollution Bulletin. 2013;76:128–38.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 591,
"serial_number": 28,
"reference": "Pierzynski GM, Sims JT, et al. Soil and environmental quality. Boca Raton, United States of America: CRC Press; 2000.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 592,
"serial_number": 29,
"reference": "Chakraborty S, Mukherjee A, et al. Biochemical characterization of a lead-tolerant strain of Aspergillus foetidus: an implication of bioremediation of lead from liquid media. International Biodeterioration and Biodegradation. 2013;84:134–42.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 593,
"serial_number": 30,
"reference": "Kabata-Pendias A, Pendias H. Trace elements in soils and plants. London: CRC Press; 2001.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 594,
"serial_number": 31,
"reference": "Perpetuo EA, Souza CB, et al. Engineering bacteria for bioremediation. In: Carpi A, editor. Progress in molecular and environmental bioengineering: from analysis and modeling to technology applications. Rijeka: InTech; 2011. p. 605–32.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 595,
"serial_number": 32,
"reference": "López-Vargas ER, Ortega-Ortíz H, et al. Foliar application of copper nanoparticles increases the fruit quality and the content of bioactive compounds in tomatoes. Applied Sciences. 2018;8(7):1020.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 596,
"serial_number": 33,
"reference": "Hamzah Saleem M, Usman K, et al. Functions and strategies for enhancing zinc availability in plants for sustainable agriculture. Frontiers in Plant Science. 2022;13:1033092.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 597,
"serial_number": 34,
"reference": "Ventrella A, Catucci L, et al. Interactions between heavy metals and photosynthetic materials studied by optical techniques. Bioelectrochemistry. 2009;77:19–25.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 598,
"serial_number": 35,
"reference": "Yan A, Wang Y, et al. Phytoremediation: a promising approach for revegetation of heavy metal-polluted land. Frontiers in Plant Science. 2020;11:359.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 599,
"serial_number": 36,
"reference": "Hassan MU, Chattha MU, et al. Nickel toxicity in plants: reasons, toxic effects, tolerance mechanisms, and remediation possibilities—a review. Environmental Science and Pollution Research. 2019;26:12673–88.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 600,
"serial_number": 37,
"reference": "Angon PB, Islam MS, Kc S, et al. Sources, effects and present perspectives of heavy metals contamination: Soil, plants and human food chain. Heliyon. 2024;10(7):e28357.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 601,
"serial_number": 38,
"reference": "Khalid S, Shahid M, et al. A comparison of technologies for remediation of heavy metal contaminated soils. Journal of Geochemical Exploration. 2017;182:247–68.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 602,
"serial_number": 39,
"reference": "Liu CJ, Deng SG, et al. Applications of bioremediation and phytoremediation in contaminated soils and waters: CREST publications during 2018–2022. Critical Reviews in Environmental Science and Technology. 2023;53:723–32.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 603,
"serial_number": 40,
"reference": "El-Sappah AH, Zhu Y, et al. Plants’ molecular behavior to heavy metals: from criticality to toxicity. Frontiers in Plant Science. 2024;15:1423625.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 604,
"serial_number": 41,
"reference": "Hama Aziz KH, Mustafa FS, et al. Heavy metal pollution in the aquatic environment: efficient and low-cost removal approaches to eliminate their toxicity: a review. RSC Advances. 2023;13:17595–17610.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 605,
"serial_number": 42,
"reference": "Abd Elnabi MK, Elkaliny NE, et al. Toxicity of heavy metals and recent advances in their removal: a review. Toxics. 2023;11:580.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 606,
"serial_number": 43,
"reference": "Podar D, Maathuis FJM. The role of roots and rhizosphere in providing tolerance to toxic metals and metalloids. Plant, Cell & Environment. 2022;45:719–36.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 607,
"serial_number": 44,
"reference": "Khan F, Siddique AB, et al. Phosphorus plays key roles in regulating plants’ physiological responses to abiotic stresses. Plants. 2023;12(15):2861.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 608,
"serial_number": 45,
"reference": "Sun S-C, Chen JX, et al. Molecular mechanisms of heavy metals resistance of Stenotrophomonas rhizophila JC1 by whole genome sequencing. Archives of Microbiology. 2021;203:2699–709.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 609,
"serial_number": 46,
"reference": "Rajput VD, Minkina T, et al. Nanotechnology in the restoration of polluted soil. Nanomaterials. 2022;12(5):769.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 610,
"serial_number": 47,
"reference": "Sharma B, Sarkar A, et al. Agricultural utilization of biosolids: a review on potential effects on soil and plant growth. Waste Management. 2017;64:117–32.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 611,
"serial_number": 48,
"reference": "Alengebawy A, Abdelkhalek ST, et al. Heavy metals and pesticides toxicity in agricultural soil and plants: ecological risks and human health implications. Toxics. 2021;9:42.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 612,
"serial_number": 49,
"reference": "Kladsomboon S, Jaiyen C, et al. Heavy metals contamination in soil, surface water, crops, and resident blood in Uthai District, Phra Nakhon Si Ayutthaya, Thailand. Environmental Geochemistry and Health. 2019;42:545–61.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 613,
"serial_number": 50,
"reference": "Singh Y, Saxena MK. Insights into the recent advances in nano-bioremediation of pesticides from the contaminated soil. Frontiers in Microbiology. 2022;13:982611.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 614,
"serial_number": 51,
"reference": "Bist P, Choudhary S. Impact of heavy metal toxicity on the gut microbiota and its relationship with metabolites and future probiotics strategy: a review. Biological Trace Element Research. 2022;200(12):5328–50.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 615,
"serial_number": 52,
"reference": "Enez A, Hudek L, et al. Reduction in trace element-mediated oxidative stress towards cropped plants via beneficial microbes in irrigated cropping systems: a review. Applied Sciences. 2018;8(10):1953.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 616,
"serial_number": 53,
"reference": "Alabresm A, Chen YP, et al. A novel method for the synergistic remediation of oil-water mixtures using nanoparticles and oil-degrading bacteria. Science of the Total Environment. 2018;630:1292–1297.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 617,
"serial_number": 54,
"reference": "Rajput VD, Kumari S, Minkina T, Sushkova S, Mandzhieva S. Nano-enhanced microbial remediation of PAHs contaminated soil. Air, Soil and Water Research. 2023;16:1–7.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 618,
"serial_number": 55,
"reference": "Cao X, Alabresm A, et al. Improved metal remediation using a combined bacterial and nanoscience approach. Science of the Total Environment. 2020;704:135378.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 619,
"serial_number": 56,
"reference": "Pandit C, Roy A, et al. Biological agents for synthesis of nanoparticles and their applications. Journal of King Saud University – Science. 2022;34(3):101869.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 620,
"serial_number": 57,
"reference": "Mane PC, Bhosle AB. Bioremoval of some metals by living algae Spirogyra sp. and Spirullina sp. from aqueous solution. International Journal of Environmental Research. 2012;6(2):571–6.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 621,
"serial_number": 58,
"reference": "Bala S, Garg D, et al. Recent strategies for bioremediation of emerging pollutants: a review for a green and sustainable environment. Toxics. 2022;10:484.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 622,
"serial_number": 59,
"reference": "Hu Y, Mortimer PE, et al. Mushroom cultivation for soil amendment and bioremediation. Circular Agricultural Systems. 2021;1(1):1–14.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 623,
"serial_number": 60,
"reference": "Rasha FM. Intracellular siderophore detection in an Egyptian cobalt-treated isolate using SEM-EDX with reference to its tolerance. Polish Journal of Microbiology. 2017;66(2):235–43.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 624,
"serial_number": 61,
"reference": "Singh MP, Vishwakarma SK, et al. Bioremediation of direct blue 14 and extracellular ligninolytic enzyme production by white rot fungi: Pleurotus spp. BioMed Research International. 2013;2013:180156.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 625,
"serial_number": 62,
"reference": "Armendariz AL, Talano MA, et al. Impact of double inoculation with Bradyrhizobium japonicum E109 and Azospirillum brasilense Az39 on soybean plants grown under arsenic stress. Plant Physiology and Biochemistry. 2019; 138:26–35.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 626,
"serial_number": 63,
"reference": "Bruno LB, Anbuganesan V, et al. Enhanced phytoextraction of multi-metal contaminated soils under increased atmospheric temperature by bioaugmentation with plant growth-promoting Bacillus cereus. Journal of Environmental Management. 2021; 289:112553.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 627,
"serial_number": 64,
"reference": "Kumar A, Voropaeva O, et al. Bioaugmentation with copper-tolerant endophyte Pseudomonas lurida strain EOO26 for improved plant growth and copper phytoremediation by Helianthus annuus. Chemosphere. 2021; 266:128983.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 628,
"serial_number": 65,
"reference": "Ahluwalia SS, Goyal D. Microbial and plant-derived biomass for removal of heavy metals from wastewater. Bioresource Technology. 2007;98(12):2243–2257.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 629,
"serial_number": 66,
"reference": "Jeyasundar PGSA, Ali A, et al. Green remediation of toxic metals contaminated mining soil using bacterial consortium and Brassica juncea. Environmental Pollution. 2021; 277:116789.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 630,
"serial_number": 67,
"reference": "Jampasri K, Pokethitiyook P, et al. Bacteria-assisted phytoremediation of fuel oil and lead co-contaminated soil in the salt-stressed condition by Chromolaena odorata and Micrococcus luteus. International Journal of Phytoremediation. 2020; 22:322–33.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 631,
"serial_number": 68,
"reference": "Yang X, Qin J, et al. Upland rice intercropping with Solanum nigrum inoculated with arbuscular mycorrhizal fungi reduces grain cadmium while promoting phytoremediation of cadmium-contaminated soil. Journal of Hazardous Materials. 2021; 406:124325.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 632,
"serial_number": 69,
"reference": "Irshad S, Xie Z, et al. Indigenous strain Bacillus XZM-assisted phytoremediation and detoxification of arsenic in Vallisneria denseserrulata. Journal of Hazardous Materials. 2020; 381:120903.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 633,
"serial_number": 70,
"reference": "Chaturvedi R, Favas PJ, et al. Harnessing Pisum sativum-Glomus mosseae symbiosis for phytoremediation of soil contaminated with lead, cadmium, and arsenic. International Journal of Phytoremediation. 2021; 23:279–90.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 634,
"serial_number": 71,
"reference": "Machado MD, Soares EV, et al. Removal of heavy metals using a brewer’s yeast strain of Saccharomyces cerevisiae: chemical speciation as a tool in the prediction and improving of treatment efficiency of real electroplating effluents. Journal of Hazardous Materials. 2010;180(1–3):347–53.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 635,
"serial_number": 72,
"reference": "Kullu B, Patra DK, et al. AM fungi-mediated bioaccumulation of hexavalent chromium in Brachiaria mutica: A mycorrhizal phytoremediation approach. Chemosphere. 2020; 258:127337.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 636,
"serial_number": 73,
"reference": "Lee YC, Chang SP. The biosorption of heavy metals from aqueous solution by Spirogyra and Cladophora filamentous macroalgae. Bioresource Technology. 2011;102(9):5297–304.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 637,
"serial_number": 74,
"reference": "Manyangadze M, Chikuruwo NHM, et al. Enhancing adsorption capacity of nano-adsorbents via surface modification: a review. South African Journal of Chemical Engineering. 2020;31(1):25–32.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 638,
"serial_number": 75,
"reference": "Tripathi P, Khare P, et al. Bioremediation of arsenic by soil methylating fungi: role of Humicola sp. strain 2WS1 in amelioration of arsenic phytotoxicity in Bacopa monnieri L. Science of the Total Environment. 2020; 716:136758.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 639,
"serial_number": 76,
"reference": "Saunders RJ, Paul NA, et al. Sustainable sources of biomass for bioremediation of heavy metals in wastewater derived from coal-fired power generation. PLoS ONE. 2012;7(5):e36470.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 640,
"serial_number": 77,
"reference": "Cantamessa S, Massa N, et al. Phytoremediation of a highly arsenic-polluted site, using Pteris vittata L. and arbuscular mycorrhizal fungi. Plant Theory. 2020; 9:1211.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 641,
"serial_number": 78,
"reference": "Saeed-ur-Rahman MK, Kayani SI, et al. The ameliorative effects of exogenous inoculation of Piriformospora indica on molecular, biochemical, and physiological parameters of Artemisia annua L. under arsenic stress condition. Ecotoxicology and Environmental Safety. 2020; 206:111202.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 642,
"serial_number": 79,
"reference": "You Y, Wang L, et al. Effects of arbuscular mycorrhizal fungi on the growth and toxic element uptake of Phragmites australis under zinc/cadmium stress. Ecotoxicology and Environmental Safety. 2021; 213:112023.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 643,
"serial_number": 80,
"reference": "Złoch M, Kowalkowski T, et al. Modeling of phytoextraction efficiency of microbially stimulated Salix dasyclados L. in soils with different speciation of heavy metals. International Journal of Phytoremediation. 2017; 19:1150–64.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 644,
"serial_number": 81,
"reference": "Fan M, Xiao X, et al. Enhanced phytoremediation of Robinia pseudoacacia in heavy metal-contaminated soils with rhizobia and the associated bacterial community structure and function. Chemosphere. 2018; 197:729–40.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 645,
"serial_number": 82,
"reference": "Tiwari N, Santhiya D, et al. Microbial remediation of micro-nano plastics: current knowledge and future trends. Environmental Pollution. 2020;265:115044.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 646,
"serial_number": 83,
"reference": "Rogers KL, Carreres-Calabuig JA, et al. Micro-by-micro interactions: how microorganisms influence the fate of marine microplastics. Limnology and Oceanography. 2020;5:18–36.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 647,
"serial_number": 84,
"reference": "Othman AR, Hasan HA, et al. Microbial degradation of microplastics by enzymatic processes: a review. Environmental Chemistry Letters. 2021;1–17.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 648,
"serial_number": 85,
"reference": "Knott BC, Erickson MD, et al. Characterization and engineering of a two-enzyme system for plastics depolymerization. Proceedings of the National Academy of Sciences. 2020;117(41):25476–85.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 649,
"serial_number": 86,
"reference": "Anastopoulos I, Pashalidis I. Single-use surgical face masks, as a potential source of microplastics: do they act as pollutant carriers?. Journal of Molecular Liquids. 2021;326:115247.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 650,
"serial_number": 87,
"reference": "Golwala H, Zhang X, et al. Solid waste: an overlooked source of microplastics to the environment. Science of the Total Environment. 2021;144581.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 651,
"serial_number": 88,
"reference": "Ru J, Huo Y, et al. Microbial degradation and valorization of plastic wastes. Frontiers in Microbiology. 2020;11:442.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 652,
"serial_number": 89,
"reference": "Beddow J, Stolpe B, et al. Effects of engineered silver nanoparticles on the growth and activity of ecologically important microbes. Environmental Microbiology Reports. 2014;6(5):448–58.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 653,
"serial_number": 90,
"reference": "Saharan BS, Chaudhary T, Mandal BS, Kumar D, Kumar R, Sadh PK, Duhan JS. Microbe-plant interactions targeting metal stress: new dimensions for bioremediation applications. Journal of Xenobiotics. 2023;13(2):252-269.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 654,
"serial_number": 91,
"reference": "Manganyi MC, Dikobe TB, Maseme MR. Exploring the potential of endophytic microorganisms and nanoparticles for enhanced water remediation. Molecules. 2024;29(12):2858.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 655,
"serial_number": 92,
"reference": "Ahmed T, Noman M, et al. Dynamic interplay between nano-enabled agrochemicals and the plant-associated microbiome. Trends in Plant Science. 2023;28(11):1310–25.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 656,
"serial_number": 93,
"reference": "Masud MAA, Shin WS. Advanced carbo-catalytic degradation of antibiotics using conductive polymer-seaweed biochar composite: exploring N/S functionalization and non-radical dynamics. Journal of Hazardous Materials. 2024;478:135449.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 657,
"serial_number": 94,
"reference": "Wang D, Pan Q, et al. Effects of mixtures of engineered nanoparticles and cocontaminants on anaerobic digestion. Environmental Science & Technology. 2024;58(6):2598–614.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 658,
"serial_number": 95,
"reference": "Shemawar, Mahmood A, Hussain S, Mahmood F, Iqbal M, Shahid M, Ibrahim M, Ali MA, Shahzad T. Toxicity of biogenic zinc oxide nanoparticles to soil organic matter cycling and their interaction with rice-straw derived biochar. Scientific Reports. 2021;11(1):8429.",
"pmc": null,
"DOI": null,
"article": 16
},
{
"id": 659,
"serial_number": 96,
"reference": "Singh Y, Saxena MK. Insights into the recent advances in nano-bioremediation of pesticides from the contaminated soil. Frontiers in Microbiology. 2022;13:982611.",
"pmc": null,
"DOI": null,
"article": 16
}
]
},
{
"id": 14,
"slug": "280-1722660642",
"featured": false,
"slider": false,
"issue": "Vol2 Issue4",
"type": "review_article",
"manuscript_id": "280-1722660642",
"recieved": "2024-08-12",
"revised": null,
"accepted": "2024-10-15",
"published": "2024-12-06",
"pdf_file": "https://plant-trends.bsmiab.org/media/pdf_file/2024/48/280-1722660642.pdf",
"title": "Exploring the cancer-fighting properties of <span>Catharanthus roseus</span>: A natural powerhouse",
"abstract": "<p><em>Catharanthus roseus</em> contains high-value medicinal properties, which are widely used in treating diabetes, kidney, liver, cardiovascular, and other life-threatening diseases. This study aims to explore the potential properties of <em>C. roseus</em> that are used as cancer-fighting agents and update the prospects and pharmacological significance of <em>C. roseus</em>. This study highlights the screening of anticancer properties, <em>in vitro</em> phytochemicals, and pharmacological advancements through biological cell culture approaches. The updated findings further suggest the potential pathways of inhibiting cancer cells and prospect of the medicinal properties in healing life-threatening diseases. The findings together open new insights into using C.<em> roseus</em> properties in pharmaceutical significance, cancer treatment, and patient care.</p>",
"journal_reference": "Plant Trends. 2024; 2(4): 108-115",
"academic_editor": "Md Jamal Uddin, PhD; ABEx Bio-Research Center, Dhaka Bangladesh",
"cite_info": "Sabrina S, Saha SR, et al. Exploring the cancer-fighting properties of Catharanthus roseus: A natural powerhouse. Plant Trends. 2024; 2(4): 108-115.",
"keywords": [
"Pollution",
"Powerhouse",
"Catharanthus roseus",
"Phytochemicals",
"Cancer"
],
"DOI": "10.5455/pt.2024.09",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>Cancer is a life-threatening disease caused by abnormal rapid cell division in a part or organ of the body. The primary cause of mortality from cancer is metastasis, where the abnormal cells rapidly increase in the body [<a href=\"#r-1\">1</a>]. According to the World Health Organization (WHO), cancer is the second leading cause of death globally, accounting for an estimated 9.6 million deaths, or 1 in 6 deaths, in 2018 [<a href=\"#r-2\">2</a>]. The American Cancer Society (ACS) published a Global Cancer Statistics report in 2024. The report revealed that approximately 20 million new cancer cases were diagnosed globally in 2022, and the disease claimed the lives of 9.7 million people worldwide [<a href=\"#r-3\">3</a>]. As cancer detection and treatment improve in high-income countries, it is expected that over the next decade, more than 75% of cancer-related deaths will occur in low- and middle-income countries [<a href=\"#r-4\">4</a>].</p>\r\n\r\n<p><em>Catharanthus roseus</em>, commonly known as Madagascar periwinkle, is a perennial plant from the Apocynaceae family [<a href=\"#r-5\">5, 6</a>]. The plant’s name is derived from Greek and means “pure flower.” This plant thrives in tropical regions and is cultivated both as an ornamental and medicinal plant. The plant extracts potential properties of <em>C. roseus</em> demonstrate a variety of pharmacological activities being used as anticancer, antidiabetic, antimicrobial, antioxidant, and other patient cures [<a href=\"#r-7\">7</a>]. <em>C. roseus</em> synthesizes more than 120 alkaloids, with 70 exhibiting pharmacological activity, including indole alkaloids such as ajmalicine, serpentine, and reserpine [<a href=\"#r-6\">6</a>]. These alkaloids possess anti-hypertensive and antispasmodic properties, which are valuable in clinical treatments. Plant compounds play showed defense against microbial infections and environmental stressors like UV radiation [<a href=\"#r-6\">6</a>].</p>\r\n\r\n<p>Among the alkaloids found in <em>C. roseus</em>, vincristine, and vinblastine are particularly notable for their anticancer properties [<a href=\"#r-8\">8</a>]. These compounds, present in the plant’s leaves, inhibit tumor growth by preventing cell division. Vincristine and vinblastine achieve this by binding to tubulin, a fundamental protein in the cytoplasm, thereby suppressing the formation of microtubule structures. These alkaloids are commonly prescribed to treat various cancers, including leukemia, Hodgkin’s lymphoma, and breast cancer [<a href=\"#r-8\">8, 9</a>].</p>\r\n\r\n<p> It is not clear how <em>C. roseus</em> alkaloids interact with cancer cells at the molecular level. Recent studies have begun to explore tentative pathways, but comprehensive insights are still lacking. A study highlighted newly isolated indole alkaloids from <em>C. roseus</em>, such as catharoseumine and 14′,15′-didehydrocyclovinblastine, which have shown effective inhibition of human cancer cell lines <em>in vitro</em> [<a href=\"#r-10\">10</a>]. Another research assessed the anticancer effects of incensole acetate, a terpenoid molecule isolated from <em>C. roseus</em> essential oil, using <em>in silico</em> docking studies and <em>in vivo</em> models [<a href=\"#r-11\">11</a>]. Nevertheless, a study explored the regulation of genes coding for enzymes involved in the synthesis of terpenoid indole alkaloids (TIA) and elucidated some molecular mechanisms controlling gene expression in cell suspension cultures of <em>C. roseus </em>[<a href=\"#r-6\">6</a>].</p>\r\n\r\n<p>Potential clinical trials are required to validate the efficacy and safety of <em>C. roseus</em>-derived compounds in diverse patients. A study emphasized the importance of clinical trials for these compounds, which suggests their potential in cancer treatment [<a href=\"#r-12\">12</a>]. Research highlighted the need for improved formulations to enhance the bioavailability and pharmacokinetics of these compounds [<a href=\"#r-9\">9</a>]. This is crucial to explore their therapeutic potential. The efficiency of the compounds has been identified as their role in resistance mechanisms in cancer cells [<a href=\"#r-13\">13</a>], but further research is needed to develop strategies to overcome this resistance. In contrast, it has been studied that cancer cells can develop resistance to alkaloids [<a href=\"#r-9\">9</a>]. </p>\r\n\r\n<p>Moreover, the synergistic effects of combining these alkaloids with other treatments suggested that combining <em>C. roseus</em> alkaloids with other therapeutic agents could enhance efficacy, but it is necessary to clarify optimal doses for therapeutic [<a href=\"#r-12\">12</a>]. <em>C. roseus</em> produces a unique set of alkaloids, including vinblastine and vincristine, which have been used in cancer treatment. A recent study identified new alkaloids such as catharoseumine and 14′,15′-didehydrocyclovinblastine, which exhibit significant anti-cancer properties [<a href=\"#r-10\">10</a>]. These compounds show potential therapeutic benefits, particularly in inhibiting the growth of cancer cell lines.</p>\r\n\r\n<p>Pham <em>et al. </em>also<em> </em>provided deeper insights into three mechanisms of <em>C. roseus </em>alkaloids. Firstly, apoptosis induction triggers programmed cell death in cancer cells. Secondly, cell cycle arrest halts the proliferation of cancer cells at specific stages of the cell cycle. (3) Inhibition of Angiogenesis: Preventing the formation of new blood vessels that supply nutrients to tumors. Compounds derived from <em>C. roseus</em> can have synergistic effects when used in combination with other cancer therapies. This synergy can enhance the efficacy of treatment protocols, potentially leading to better outcomes for patients [<a href=\"#r-10\">10</a>]. Advances in targeted delivery systems have significantly improved the precision and efficacy of <em>C. roseus</em>-derived treatments. These systems allow the delivery of alkaloids to cancer cells, minimizing side effects and enhancing therapeutic outcomes [<a href=\"#r-10\">10</a>, <a href=\"#r-14\">14</a>].</p>\r\n\r\n<p>Research from 2020 has proposed new strategies to overcome resistance mechanisms in cancer cells. These strategies involve using <em>C. roseus</em> alkaloids in combination with other agents to bypass or counteract resistance pathways, making them a promising candidate for combination therapies [<a href=\"#r-10\">10</a>].</p>\r\n\r\n<p>In order to address these research gaps and highlight the insights and prospects of the cancer-fighting properties in <em>C. roseus</em>. In this review, we focus on the isolation of cancer-fighting properties from <em>C. roseus</em>, the insights underlying the molecular mechanism of cancer cell inhibition, and use of the components as potential therapeutics and their further pharmacological significance.</p>"
},
{
"section_number": 2,
"section_title": "CANCER-FIGHTING PROPERTIES IN C. roseus",
"body": "<p>Several potential metabolites have been isolated from <em>C. roseus</em>, which are<em> </em>presented in <a href=\"#figure1\">Figure 1</a>. Vinblastine and Vincristine are the primary bioactive alkaloids derived from <em>C. roseus</em>. Vinblastine has been used in treating Hodgkin’s lymphoma, breast cancer, and testicular cancer [<a href=\"#r-12\">12</a>]. Vincristine is effective against acute lymphoblastic leukemia and Hodgkin’s disease [<a href=\"#r-15\">15</a>]. Both vinblastine and vincristine were involved in inhibiting the formation of the mitotic spindle, which is crucial for cell division [<a href=\"#r-16\">16</a>]. This action prevents cancer cells from proliferating, thereby slowing down or stopping tumor growth [<a href=\"#r-16\">16</a>].</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"481\" src=\"/media/article_images/2024/02/08/280-1722660642-Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1. </strong>The potential metabolites found in <em>Catharanthus roseus</em>. The representative figure presents a total of nine metabolites, which are involved in anticancer and antioxidant potential. This figure was drawn with the BioRender tool (<a href=\"https://www.biorender.com/\">https://www.biorender.com/</a>).</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p>Vinblastine works by binding to tubulin, a protein that is essential for microtubule formation. Microtubules are crucial for cell division as they form the mitotic spindle, which separates chromosomes during mitosis. Vinblastine prevents the formation of microtubules that lead to tubulin polymerization, thereby arresting cells in the metaphase of mitosis (<a href=\"#figure2\">Figure 2</a>). This disruption leads to cell death, particularly in rapidly dividing cancer cells. Vincristine has a similar mechanism of action to Vinblastine (<a href=\"#figure2\">Figure 2</a>). It also binds to tubulin and inhibits microtubule formation. However, Vincristine is particularly effective in treating certain types of leukemia and lymphomas. By preventing microtubule assembly, Vincristine disrupts the mitotic spindle, leading to cell cycle arrest and apoptosis (programmed cell death) in cancer cells (<a href=\"#figure2\">Figure 2</a>).</p>\r\n\r\n<p style=\"text-align:center\"><strong>Table 1. </strong>List of key cancer-fighting properties in <em>C. roseus.</em></p>\r\n\r\n<div id=\"Table-1\">\r\n<p style=\"text-align:center\"><a href=\"https://plant-trends.bsmiab.org/table/280-1722660642-table1/\">Table-1</a></p>\r\n</div>"
},
{
"section_number": 3,
"section_title": "EXTRACTION PROCESSES AND PHYSICOCHEMICAL PROPERTIES OF C. roseus",
"body": "<p>The toxic solvent wastes produced by several industrial operations have negative impacts on thyroid, hematological, and respiratory health [<a href=\"#r-22\">22</a>]. Thus, the proper extraction processes of potential compounds are highly demandable. The separation of the bioactive component using water as the solvent is an alternate technique. Nevertheless, the component must be extracted using water that is heated to a high temperature. The purity of the extract and the targeted bioactive component may both suffer from the extraction's increased temperature. Additionally, the water extraction procedure took an extended period to finish the extraction procedure [<a href=\"#r-23\">23</a>].</p>\r\n\r\n<p>Vinblastine and vincristine are the key dimeric indole alkaloids that are frequently employed in cancer treatment, are produced in low quantities by <em>C. roseus</em>. Eli Lilly originally discovered techniques for extracting and purifying a number of the alkaloids present in <em>C. roseus</em> using organic solvents [<a href=\"#r-24\">24</a>]. There are four processes in these sulphuric acid and water extraction techniques, including fractionation by partition with benzene, two chromatographic columns, and crystallization in ethanol. Extensive progress has been made related to extracting vindoline, catharanthin, and vinblastine from <em>C. roseus</em> leaves utilizing ultrasonic extraction, methanol, heat, boiling, and refluxing. The use of supercritical fluid extraction (SFE) to extract phytochemicals from <em>C. roseus</em> has also been used [<a href=\"#r-24\">24</a>].</p>"
},
{
"section_number": 4,
"section_title": "MECHANISM OF BIOACTIVE COMPOUNDS OF C. roseus AGAINST CANCER",
"body": "<p>Indole alkaloids produced by <em>C. roseus</em> are known as Vinka alkaloids and are frequently used as antimitotic medications to treat cancer. Natural compounds like vincristine and vinblastine are anticancer compounds applied in clinical trials [<a href=\"#r-25\">25</a>]. Vinca alkaloids change the kinetics of the microtubules, causing apoptosis and suppressing the development of cells. These microtubules (MTs), known as cytoskeleton elements and important parts of the spindles that separate the chromosomes amid meiosis and mitosis, are also crucial for preserving cell shape, transport, and a variety of other cellular functions [<a href=\"#r-26\">26</a>]. Tubulin and tubulin heterodimers, which dynamically polymerize and depolymerize at the edges, are the fundamental building blocks of the MTs; The interaction of tubulin and guanosine 5-triphosphate (GTP) controls the formation and breakdown of the MT polymers, also known as treadmilling and dynamic instability, and any destabilization or interruption of this interactions can stop the cell cycle and cause programmable cell death or apoptosis [<a href=\"#r-27\">27</a>]. Due to this, two categories of destabilizers have been identified. The first category includes substances that stabilize MTs and hinder depolymerization, whereas the second category includes substances that depolymerize MTs and hinder their formation [<a href=\"#r-28\">28</a>]. Vinca alkaloids and their analogy work by attaching to two tubulin heterodimers at the outermost layer near the convertible GTP-binding region and depolymerizing the MTs to stop the progression of tumor cells amid mitosis [<a href=\"#r-29\">29</a>] (<a href=\"#figure2\">Figure 2</a>). Although the apparent propensity of the two indigenous Vinca alkaloids from <em>C. roseus</em> or their partially synthetic counterparts and analogy for tubulin heterodimers is identical, it is distinguished by an additional sequence of a reduction in the overall equilibrium constants: Vincristine is followed by Vinblastine, Vinorelbine, and Vinflunine. Via shared docking premises, van der Waals forces and electrostatic interaction energy consolidate the combined structures of these alkaloids with, α, β-tubulin.</p>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"409\" src=\"/media/article_images/2024/02/08/280-1722660642-Figure2.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 2.</strong> Mechanism of action of Vinblastine and Vincristine against cancer cells. Vinblastine and 86 vincristine combat cancer by targeting microtubules within the cell. They bind to tubulin, an 87 protein that forms microtubules, disrupting their dynamics. This interference halts cell division, 88 leading to mitotic arrest and ultimately causing cancer cell death. The chemical structures are adapted from the National Center of Biotechnology Information (<a href=\"https://pubchem.ncbi.nlm.nih.gov/\">https://pubchem.ncbi.nlm.nih.gov/</a>). </figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p>Additionally, the two fluorine atoms that distinguish vinflunine from vinorelbine increase this electrostatic connection in the context of vinflunine [<a href=\"#r-30\">30</a>], the vindoline region then ensures tubulin heterodimers' interaction, as the catharanthine region provides the cytotoxic impact [<a href=\"#r-22\">22</a>]. According to some publications, vinca alkaloids work in a dose-dependent manner, preventing cell division when present in low concentrations. The cells then perish over an extended period of incubation. Nevertheless, at high concentrations, vinca alkaloids cause cell death by causing the production of paracristals (giant tubulin polymers), which prevent tumoral cells from undergoing mitosis [<a href=\"#r-31\">31</a>]. Numerous scientists have lately reported on new modes of action for these alkaloids, which include interactions with MTs-associated proteins, calmodulin, and the inhibition of amino acid metabolism [<a href=\"#r-32\">32</a>]. Using this novel method of action, nevertheless, variations in Vinca alkaloids' potency were found. Then, although having a lesser affinity for tubulin than vinblastine or vincristine, vinflunine's superior activity against murine tumors and human tumor xenografts can be explained by its interaction with calmodulin. The monoterpenoid indole alkaloid catharoseumine, which was isolated from the <em>C. roseus</em> plant, has a distinctive peroxy bridge moiety and exhibits cytotoxicity when tested on human tumor cell lines, albeit with modest cytotoxicity when tested on the HL-60 cell line.</p>"
},
{
"section_number": 5,
"section_title": "SYNTHESIS OF ANTI-CANCER AGENTS FROM C. roseus",
"body": "<p>Extensive progress has been made in the culture of plant cells for more effective synthesis of anticancer substances. These approaches open several new benefits, including the ability to produce bioactive pharmaceuticals with purification processes, a highly effective production system in short duration, and a well-controlled manufacturing process under sterile circumstances. The <em>C. roseus</em> has shown potential to produce high-value pharmaceuticals in recent years utilizing submerged growth techniques that either use shake flasks or various forms of bioreactors [<a href=\"#r-33\">33</a>]. It has been demonstrated that hairy root lines of <em>C. roseus</em> may be grown in bioreactors of various sizes for the large-scale synthesis of ajmalicine, serpentine, and catharanthine [<a href=\"#r-34\">34</a>]. Vincristine and vinblastine, two more promising anticancer bioactive agents, were effectively synthesized in stirred tank bioreactor system. These alkaloids were produced at their maximum levels when the pH was unregulated, and the aeration rate was 0.5 v/v/min. The vincristine and vinblastine were produced 13.47 and 7.94, respectively [<a href=\"#r-35\">35</a>].</p>\r\n\r\n<p>The synthesis of alkaloid compounds rises with the presence of a chemical that induce stresses to cells. According to the report, the addition of relatively small amounts of chromium to culture media (between 10 and 100 M), was found to decrease cell proliferation, while vinblastine and vincristine synthesis also significantly increased [<a href=\"#r-36\">36</a>]. The incorporation of sodium chloride into cultures to induce osmotic pressure can also result in different stress signaling, enzyme functions, and a rise in the generation of vinblastine and vincristine [<a href=\"#r-37\">37</a>]. Another study revealed the addition of the <em>Aspergillus flavus</em> fungal elicitor significantly stimulated cell proliferation and increased the synthesis of vinblastine and vincristine [<a href=\"#r-38\">38</a>]. However, one recent study showed that the supplementation of yeast extract to culture media, even at low concentrations (1.5g/L), significantly increased the levels of vinblastine and vincristine in suspension cultures of <em>C. roseus </em>[<a href=\"#r-39\">39</a>].</p>"
},
{
"section_number": 6,
"section_title": "PROSPECTS AND LIMITATIONS OF USING C. roseus",
"body": "<p>The current study confirms the extraction and isolation processes of several alkaloids including vinblastine, and vincristine, which have potent anticancer and antidiabetic properties. However, further study is required on their anticancer activities and underlying molecular mechanisms. Nanotechnology-based synthesis of these properties and formulations opens new prospects for enhancing the efficacy of cancer treatment. In contrast, toxicity concerns of the potential alkaloids and the limitations of their application should be screened properly. The extraction process of bioactive compounds should be improved, and a strategy needs to be applied to reduce pharmaceutical costs, although it remains a challenge to optimize. In addition, the alkaloid-associated side effects must be addressed properly. The solution to these limitations with the potential development of disease suppression or inhibition of cancer cell growth could open new horizons for <em>C. roseus-</em>based therapy against cancer.</p>"
},
{
"section_number": 7,
"section_title": "CONCLUSIONS",
"body": "<p>This study explored the potential vinca alkaloids found in<em> C. roseus</em>, and their prospects of use as<em> </em>anticancer compounds, and mechanical insights of vincristine and vinblastine involving inhibition of cancer cell proliferation, and the prospects of enhancing potential alkaloid synthesis using bioreactor approach. This updated study further suggests the potential alkaloids isolated from <em>C. roseus</em>, show their pharmaceutical significance with the development of <em>C. roseus</em>-derived cancer drugs. The prospect of efficient cultivation in bioreactor-based large-scale production of these potential compounds would be an excellent source of sustainable raw materials supply for the pharmaceutical industry in producing high-quality anticancer agents. Considering these potential benefits of C.<em> roseus</em>, might be considered a natural powerhouse and used as a cancer therapeutics and patient care.</p>"
},
{
"section_number": 8,
"section_title": "AUTHOR CONTRIBUTIONS",
"body": "<p>MJI and MK designed the study. SS and SRS wrote the initial draft and prepared tables and figures. MJI edited and reviewed the whole manuscript. All the authors approved the final version of the manuscript.</p>"
},
{
"section_number": 9,
"section_title": "CONFLICTS OF INTEREST",
"body": "<p>There is no conflict of interest among the authors.</p>"
}
],
"figures": [
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/02/08/280-1722660642-Figure1.jpg",
"caption": "Figure 1. The potential metabolites found in Catharanthus roseus. The representative figure presents a total of nine metabolites, which are involved in anticancer and antioxidant potential. This figure was drawn with the BioRender tool (https://www.biorender.com/).",
"featured": false
},
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/02/08/280-1722660642-Figure2.jpg",
"caption": "Figure 2. Mechanism of action of Vinblastine and Vincristine against cancer cells. Vinblastine and 86 vincristine combat cancer by targeting microtubules within the cell. They bind to tubulin, an 87 protein that forms microtubules, disrupting their dynamics. This interference halts cell division, 88 leading to mitotic arrest and ultimately causing cancer cell death. The chemical structures are adapted from the National Center of Biotechnology Information (https://pubchem.ncbi.nlm.nih.gov/).",
"featured": false
}
],
"authors": [
{
"id": 64,
"affiliation": [
{
"affiliation": "Department of Biochemistry and Molecular Biology, Mawlana Bhashani Science and Technology University, Santosh, Tangail-1902, Bangladesh"
}
],
"first_name": "Saima",
"family_name": "Sabrina",
"email": null,
"author_order": 1,
"ORCID": null,
"co_first_author": true,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 14
},
{
"id": 65,
"affiliation": [
{
"affiliation": "Department of Microbiology, Noakhali Science and Technology University, Noakhali-3814, Bangladesh"
}
],
"first_name": "Sumita Rani",
"family_name": "Saha",
"email": null,
"author_order": 2,
"ORCID": null,
"co_first_author": true,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 14
},
{
"id": 66,
"affiliation": [
{
"affiliation": "Department of Biochemistry and Molecular Biology, Mawlana Bhashani Science and Technology University, Santosh, Tangail-1902, Bangladesh"
}
],
"first_name": "Mohammad",
"family_name": "Kamruzzaman",
"email": null,
"author_order": 3,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 14
},
{
"id": 67,
"affiliation": [
{
"affiliation": "Department of Biochemistry and Molecular Biology, Mawlana Bhashani Science and Technology University, Santosh, Tangail-1902, Bangladesh"
}
],
"first_name": "Mohammod Johirul",
"family_name": "Islam",
"email": "johir7479@gmail.com",
"author_order": 4,
"ORCID": "https://orcid.org/0000-0002-4808-1685",
"co_first_author": false,
"co_author": false,
"corresponding": true,
"corresponding_author_info": "Mohammod Johirul Islam, PhD; Department of Biochemistry and Molecular Biology, Mawlana Bhashani Science and Technology University, Santosh, Tangail-1902, Bangladesh.\r\nEmail: johir7479@gmail.com",
"article": 14
}
],
"views": 443,
"downloads": 111,
"references": [
{
"id": 479,
"serial_number": 1,
"reference": "Fares J, Fares MY, et al. Molecular principles of metastasis: a hallmark of cancer revisited. Signal Transduction and Targeted Therapy. 2020 Mar 12;5(1):28.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 480,
"serial_number": 2,
"reference": "WHO. Cancer. https://www.who.int/health-topics/cancer",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 481,
"serial_number": 3,
"reference": "American Cancer Society. American Cancer Society Releases Latest Global Cancer Statistics; Cancer Cases Expected to Rise to 35 Million Worldwide by 2050. https://pressroom.cancer.org/GlobalCancerStatistics2024",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 482,
"serial_number": 4,
"reference": "Anandasabapathy S, Asirwa C, et al. Cancer burden in low-income and middle-income countries. Nat Rev Cancer. 2024 Mar;24(3):167-170.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 483,
"serial_number": 5,
"reference": "Sharma N, Singh I, et al. Catharanthus Roseus: A Source of Anticancer Phytomedicines. In: Kole, C. (eds) The Catharanthus Genome . Compendium of Plant Genomes. Springer, Cham.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 484,
"serial_number": 6,
"reference": "Jair Barrales-Cureño H, Reyes Reyes C, et al. Alkaloids of Pharmacological Importance in Catharanthus roseus.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 485,
"serial_number": 7,
"reference": "Kumar S, Singh B, et al. A review of its ethnobotany, phytochemistry, ethnopharmacology and toxicities. J Ethnopharmacol. 2022",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 486,
"serial_number": 8,
"reference": "Alam MM, Naeem M, et al. Vincristine and Vinblastine Anticancer Catharanthus Alkaloids: Pharmacological Applications and Strategies for Yield Improvement.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 487,
"serial_number": 9,
"reference": "Dhyani P, Quispe C, et al. Anticancer potential of alkaloids: a key emphasis to colchicine, vinblastine, vincristine, vindesine, vinorelbine, and vincamine. Cancer Cell Int. 2022 Jun 2;22(1):206.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 488,
"serial_number": 10,
"reference": "Pham HNT,Vuong QV, et al. Phytochemicals Derived from Catharanthus roseus and Their Health Benefits. Technologies 2020, 8, 80.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 489,
"serial_number": 11,
"reference": "Nayila I, Sharif S et al. Characterization and anti-breast cancer potential of an incensole acetate nanoemulsion from Catharanthus roseus essential oil; in silico, in vitro, and in vivo study. RSC Adv. 2023 Nov 2;13(46):32335-32362.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 490,
"serial_number": 12,
"reference": "Sathya Prabhu D, Devi Rajeswari V. Catharanthus roseus: The Cancer-Fighting Medicine. In: Naeem, M., Aftab, T., Khan, M. (eds) Catharanthus roseus. Springer, 2017.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 491,
"serial_number": 13,
"reference": "Emran TB, Shahriar A et al. Multidrug Resistance in Cancer: Understanding Molecular Mechanisms, Immunoprevention and Therapeutic Approaches. Front Oncol. 2022",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 492,
"serial_number": 14,
"reference": "Sharifi-Azad M, Fathi M, et al. Recent advances in targeted drug delivery systems for resistant colorectal cancer. Cancer Cell Int. 2022 May 19;22(1):196.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 493,
"serial_number": 15,
"reference": "Newitt J. Catharanthus roseus: A cancer-killing champion of the plant world. 2018. https://www.kew.org/read-and-watch/catharanthus-roseus-cancer-killing-plant",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 494,
"serial_number": 16,
"reference": "Naeem, M. & Aftab et al. (2017). Catharanthus roseus: Current Research and Future Prospects.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 495,
"serial_number": 17,
"reference": "Nguyen DQ, Nguyen NL,et al . Isolation and Identification of Vincristine and Vinblastine Producing Endophytic Fungi from Catharanthus roseus (L.) G. Don. Russ J Plant Physiol 70, 188 (2023).",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 496,
"serial_number": 18,
"reference": "Koel, Mihkel & Kuhtinskaja, et al(2020). Extraction of Bioactive Compounds from Catharanthus roseus and Vinca minor. Separation and Purification Technology.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 497,
"serial_number": 19,
"reference": "Amini, Younes & Hassanvand,et al. Optimization of liquid-liquid extraction of calcium with a serpentine microfluidic device. International Communications in Heat and Mass Transfer. 140. 106551.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 498,
"serial_number": 20,
"reference": "Asia, Sammer, Y. et al. Structure and Synthesis of Vindolicine and Derivatives. Chemistry & biodiversity, 21(4).",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 499,
"serial_number": 21,
"reference": "Siddharth Goswami, Amena Ali,et al Madhulika Esther Prasad, Pallavi Singh, Pharmacological significance of Catharanthus roseus in cancer management: A review, Pharmacological Research Modern",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 500,
"serial_number": 22,
"reference": "Babu NS, Reddy SM. 2014. Impact of solvents leading to environmental pollution. Journal of Chemical and Pharmaceutical Sciences. Special issue 3, 132-35.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 501,
"serial_number": 23,
"reference": "Wojdylo A, Oszmiański J, et al. Composition and quantification of major polyphenolic compounds, antioxidant activity and colour properties of quince and mixed quince jams. Int. J. Food Sciences and Nutrition. 64(6): 749-56.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 502,
"serial_number": 24,
"reference": "Falcao MA, Scopel R,et al. Supercritical fluid extraction of vinblastine from Catharanthus roseus. The Journal of Supercritical Fluids. 129: 9-15.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 503,
"serial_number": 25,
"reference": "Sottomayor M, Ros Barcelo A. 2005.The Vinca alkaloids: from biosynthesis and accumulation in plant cells, to uptake, activity and metabolism in animal cells. In: Attaur R., editor. Studies in natural products chemistry (Bioactive natural products). Amsterdam: Elsevier Science Publisher. 813-57.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 504,
"serial_number": 26,
"reference": "Wang L., He HP, et al. Catharoseumine, a new monoterpenoid indole alkaloid possessing a peroxy bridge from Catharanthus roseus. Tetrahedron Letters. 53(13), 1576-1578.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 505,
"serial_number": 27,
"reference": "Jordan MA. 2002. Mechanism of action of antitumor drugs that interact with microtubules and tubulin. Current Medicinal Chemistry-AntiCancer Agents. 2(1): 1-17.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 506,
"serial_number": 28,
"reference": "Perez EA. 2009. Microtubule inhibitors: Differentiating tubulin-inhibiting agents based on mechanisms of action, clinical activity, and resistance. Molecular Cancer Therapeutics. 8: 2086-95.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 507,
"serial_number": 29,
"reference": "Bolanos-Garcia VM. 2009. Assessment of the mitotic spindle assembly checkpoint (SAC) as the target of anticancer therapies. Current Cancer Drug Targets. 9(2): 131-41.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 508,
"serial_number": 30,
"reference": "Coderch C, Morreale A, et al. Tubulinbased structure-affinity relationships for antimitotic Vinca alkaloids. Anti-Cancer Agents in Medicinal Chemistry (Formerly Current Medicinal Chemistry-Anti-Cancer Agents). 12(3): 219-25.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 509,
"serial_number": 31,
"reference": "Attard G, Greystoke A, et al. Update on tubulin-binding agents. Pathologie Biologie. 54(2): 72-84.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 510,
"serial_number": 32,
"reference": "Barbier P., Tsvetkov PO, et al. Deciphering the molecular mechanisms of anti-tubulin plant derived drugs. Phytochemistry Reviews. 13(1): 157-169.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 511,
"serial_number": 33,
"reference": "Mujib A, Ali M, et al2014. Somatic embryo mediated mass production of Catharanthus roseus in culture vessel (bioreactor) – A comparative study. Saudi Journal of Biological Sciences. 21: 442-49.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 512,
"serial_number": 34,
"reference": "Verma P, Mathur AK. and Shanker K. 2012. Growth, alkaloid production, rol genes integration, bioreactor up-scaling and plant regeneration studies in hairy root lines of Cathranthus roseus. Plant Biosystems – An International Journal Dealing with all Aspects of Plant Biology. 146: 27-40.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 513,
"serial_number": 35,
"reference": "Taha HS, Shamas KA, Nazif NM, et al. In vitro studies on Egyptian C. roseus (L.) G.Don V: Impact of stirred reactor physical factors on achievement of cells proliferation and vincristine and vinblastine accumulation. Research J. Pharmaceutical, Biological and Chemical Sciences. 5: 330-40.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 514,
"serial_number": 36,
"reference": "Rai V., Tandon PK. and Khaltoon S. 2014. Effect of chromium on antioxidant potential of Catharanthus roseus varieties and production of their anticancer alkaloids: vincristine and vinblastine. BioMed Research International. Vol2014, Article ID 934182, 10p.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 515,
"serial_number": 37,
"reference": "Fatima S., Mujib A. and Tonk D. 2015. NaCl amendment improves vinblastine and vincristine synthesis in Catharanthus roseus: a case of stress signalling as evidence by antioxidant enzymes activities. Plant Cell, Tissue and Organ Culture (PCTOC). 121(2): 445-458.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 516,
"serial_number": 38,
"reference": "Tonk D, Mujin A, et al. Aspergillus flavus fungus elicitation improves vincristine and vinblastine yield by augmenting callus biomass growth in Catharanthus roseus. Plant Cell, Tissue and Organ Culture (PCTOC). 126(2): 291-303.",
"pmc": null,
"DOI": null,
"article": 14
},
{
"id": 517,
"serial_number": 39,
"reference": "Maqsood M. and Abdul M. 2017. Yeast extract elicitation increases vinblastine and vincristine yield in protoplast derived tissues and plantlets in Catharanthus roseus. Revista Brasileira de Farmacognosia. 27(5): 549-56.",
"pmc": null,
"DOI": null,
"article": 14
}
]
},
{
"id": 15,
"slug": "280-1722352436",
"featured": true,
"slider": true,
"issue": "Vol2 Issue4",
"type": "original_article",
"manuscript_id": "280-1722352436",
"recieved": "2024-07-30",
"revised": null,
"accepted": "2024-11-16",
"published": "2024-12-05",
"pdf_file": "https://plant-trends.bsmiab.org/media/pdf_file/2024/28/280-1722352436.pdf",
"title": "Utilization of endophytic bacteria for liquid biofertilizer production with a newly designed prototype fermentor for plant improvement",
"abstract": "<p>Endophytic bacteria are a promising source of cost-effective and eco-friendly approaches that are highly desirable for sustainable agriculture. The study successfully developed a novel prototype fermentor and demonstrated an impactful utilization of endophytic bacteria for large-scale production of biofertilizers. The fermentor was designed to integrate i) a temperature control unit, ii) a heating unit, a heated water circulation unit, and iii) a microbial growth unit, ensuring precise control of temperature and agitation, providing a conducive environment containing an affordable medium for the proliferation of single endophytic bacterium, or consortium of endophytic bacteria. The capacities of the fermentor revealed that the temperature of the growth medium could be elevated from 29°C to 37°C within 50 minutes at varying rotation speeds of 60 rpm, 75 rpm, or 90 rpm, securing uniform temperature distribution for optimum growth of the consortia. Each consortium's average growth rate was recorded after 48 hours. Furthermore, the system efficiently returned to the initial temperature of 29°C from 37°C within 4.5 hours and maintained a temperature around 35°C to 37°C during a brief period of over ~70 minutes of power outage. The amplified culture of consortium A. comprised of growth-promoting endophytic bacteria (<em>Klebsiella </em>sp. HSTU-Bk11, <em>Acinetobacter</em> sp. HSTU-Abk29, <em>Citrobacter</em> sp. HSTU-ABk30, and <em>Enterobacter cloacae</em> HSTU-ABk39) significantly improved the morphological traits of rice plants (root, shoot, tillers number) along with the crop yield in the fields. This study presents a novel fermentor, facilitating the large-scale production of plant growth-promoting endophytic bacteria, and presenting new insights for enhancing sustainable organic agricultural research and industrial applications.</p>",
"journal_reference": "Plant Trends. 2024; 2(4): 92-107.",
"academic_editor": "Izhar Ali, PhD, Guangxi University, China",
"cite_info": "Ahmed T, Rahman MS, et al., Utilization of endophytic bacteria for liquid biofertilizer production with a newly designed prototype fermentor for plant improvement. Plant Trends. 2024 Dec; 2(4): 92-107.",
"keywords": [
"endophytic bacteria",
"Microbial growth",
"biofertilizer",
"sustainable agriculture",
"Fermentor"
],
"DOI": "10.5455/pt.2024.08",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>The world's population is projected to reach 9.7 billion by the year 2050 and 11 billion by the year 2100 [<a href=\"#r-1\">1</a>] with an annual growth rate of 0.88% [<a href=\"#r-2\">2</a>], making access to healthy nutrition unreachable, raising a critical concern. The global demand for food is predicted to increase by 35% to 56% from 2010 to 2050, while the proportion of people at risk of hunger is expected to shift from a negative 91% to a positive 8% during the same period [<a href=\"#r-3\">3</a>, <a href=\"#r-4\">4</a>]. To augment the agricultural output to meet the increased demand along with the scarcity of available lands, farmers faced critical choices, including the use of chemical fertilizers/pesticides more and more to increase crop production. The excessive applications of chemical fertilizers over time degrade soil properties and lower soil fertility [<a href=\"#r-5\">5</a>]; increase water pollution, and alter soil pH acidity, causing a decrease in the microbial population in the rhizosphere and a rise in the vulnerability of the crops to insects [<a href=\"#r-6\">6</a>]. Furthermore, rice production in Bangladesh heavily depends on the use of fertilizers with an average application rate of over 286 kg/hectare. The country imports over 1.2 million tons of chemical fertilizers every year, meeting the needs of 31%, 57%, and 95% of nitrogen, phosphate, and potash, respectively [<a href=\"#r-7\">7</a>], and impacting the foreign reserves. On the other hand, organic fertilizers enhance the soil's fertility, physical and chemical properties, ability to retain water, fungal-to-bacterial ratio, and soil enzyme activity, hence crop yields [<a href=\"#r-8\">8</a>]. The efficient use of plant growth-promoting rhizobacteria (PGPR) in agriculture has been demonstrated to replace the application of chemical fertilizers and subsequently help the development of eco-friendly crop production [<a href=\"#r-9\">9</a>, <a href=\"#r-10\">10</a>]. Furthermore, plant growth, root development patterns, competitiveness in obtaining nutrition from the soil, and defense against biotic and abiotic stresses can be improved by plant growth-promoting rhizobacteria (PGPR) application [<a href=\"#r-10\">10-13</a>]. Many rhizospheric bacteria, including <em>Azospirillum, Bacillus, Pseudomonas</em>, and <em>Enterobacter</em> have been employed to increase crop production under various stress conditions [<a href=\"#r-14\">14-16</a>]. Rhizospheric bacteria, such as <em>Pseudomonas </em>sp., <em>Azospirillum</em> sp., <em>Pantoea</em> sp., and <em>Agrobacterium </em>sp. enhanced nutrient absorption under mild temperatures by several crops, including maize, wheat, and legumes, and subsequent yields [<a href=\"#r-17\">17</a>]. These PGPRs not only have the capacity to colonize plant roots, benefiting their hosts by regulating phytohormone synthesis, enhancing the availability of soil minerals, and enhancing disease resistance, but also can act as phytostimulators, changing the metabolism of phytohormones by increasing the synthesis of auxin, cytokinins, abscisic acid, gibberellins, and by reducing ethylene [<a href=\"#r-18\">18-20</a>]. In addition to reducing the use of chemical fertilizers, PGPR has been reported to enhance crop production by effectively mitigating biotic and abiotic factors and abiotic stresses [<a href=\"#r-21\">21-23</a>]. Through competition for nutrients, antagonistic relationships, and the induction of systemic resistance, PGPR also indirectly serves as biopesticides or biocontrol agents, enhancing resistance to phytopathogens [<a href=\"#r-18\">18</a>, <a href=\"#r-19\">19</a>, <a href=\"#r-23\">23</a>].</p>\r\n\r\n<p>Under ideal conditions, a bacterium growing on a homogenous rich culture medium can multiply in as little as twenty minutes [<a href=\"#r-24\">24</a>]. On the other hand, it has been proposed that in some deep terrestrial habitats, cell division might happen as slowly as once per 100 years [<a href=\"#r-25\">25</a>] and has been attributed to the numerous variables, including nutrient-poor subsurface ecosystems responsible for the delayed development of cells. Consequently, cells are more likely to remain isolated, unable to exchange nutrients or activate defense systems, and grow more slowly [<a href=\"#r-25\">25</a>]. Thus, a system enabling regulated changes with desired precision under various circumstances is essential for the large-scale production of bacteria to be used as bio-fertilizers. Furthermore, the system a fermentor, provides a safe and repeatable technological resource required to conduct carefully thought-out studies aimed at understanding the biological, chemical, and/or physical effects on the uniform growth of the bacterial consortium. An effective fermentor offers advantages in terms of minimal risk of contamination, simplicity of use, and scalability for eventual applications in the fields. To date, due to the design complexity of the fermentor, the output objectives and expectations in the process of developing the system have only been met partially [<a href=\"#r-26\">26</a>]. To promote optimal bacterial growth, a regulated microenvironment with the ability of precise mechanical stimulation is essential. Through effective mixing, ideal mass, and heat transmission, along with a regulated growth environment, such as temperature, the fermentor is intended to do its purpose efficiently, providing an ideal condition for enhancing bacterial culture. To prevent inappropriate mixing, resulting in a heterogeneous environment and inhibition of growth [<a href=\"#r-27\">27</a>], the fermentor should be simple to scale up the production of biofertilizers. However, scaling up biofertilizer production presents multiple challenges, such as achieving high cell concentrations, ensuring product safety by removing contaminants, and lowering production costs [<a href=\"#r-28\">28</a>]. On the other hand, the production of biofertilizers (consortia of endophytic bacteria) can be costly due to the expensive media along with the available fermentors with limited capacity to culture the bacterial consortium. Traditional media used for large-scale cultures of bacterial biofertilizer is typically made from expensive ingredients, such as peptone and yeast extract. Hence, we have developed a new inexpensive growth medium and designed a new fermentor, demonstrable for large-scale bacterial cultures, meeting the needs of next-generation biofertilizers for sustainable, cost-effective, and environmentally friendly organic agriculture. The medium is made from readily available cheap ingredients, such as poultry feed, maize flour, and wheat flour.</p>\r\n\r\n<p>We have previously reported that the yield of rice grains was increased by 30-35% with a foliar spray of endophytic consortia [<a href=\"#r-29\">29, 30</a>] and the production of tomatoes was increased by 2.5 folds with a foliar spray of <em>Enterobacter</em> sp. strain HSTU-Sh6 [<a href=\"#r-30\">30</a>]. However, the production scale of the biofertilizers to cover the large area of farmlands was limited due to the use of the flask culture method. At present, the large-scale production of biofertilizers is greatly lacking in Bangladesh and worldwide due to a lack of needed technologies. This demands to development of a new cost-effective commercial fermentor to scale up the production of liquid-based endophytic bacterial consortia as biofertilizers. To this effect, the newly invented prototype fermentor reported here has the potential to revolutionize the production of biofertilizers, making it more affordable and accessible to farmers worldwide. Our group built the new fermentor in the consortium at a low cost with individual temperature, and agitation, and is suitable for use in undeveloped areas where outage of electricity is frequent.</p>"
},
{
"section_number": 2,
"section_title": "MATERIALS AND METHODS",
"body": "<p><strong>Design and fabrication of a new fermentor</strong></p>\r\n\r\n<p>A fermentor was newly designed comprised of multiple components, and is capable of producing repeatable outcomes with precision. These components were made from different materials with various shapes and sizes, depending on the specific requirements of the design. Different parts were assembled by cutting, drilling, and welding, which was required based on the availability of the resources [<a href=\"#r-27\">27</a>]. For our prototype, we chose a 2 mm sheet of stainless steel to construct the fermentor because of its lasting resistance to rust, corrosion, and discoloration, maintaining a hygiene and sanitary milieu essential for bacterial growth and culture [<a href=\"#r-31\">31</a>]. In addition, the flat, non-porous surface of stainless steel is naturally resistant to bacterial adhesion, making it simpler to manage and monitor bacterial proliferation without interference from the material [<a href=\"#r-32\">32</a>]. Overall, these features of stainless steel are especially important in maintaining integrity, producing reliable results, and reducing the risk of contamination. The fermentor (<a href=\"#figure1\">Figure 1</a>) was introduced includes a power source, an inner chamber defined as a sample enclosure, and an outer chamber. An access is included through which a test sample is placed within the inner chamber. The outer chamber wraps around one or more walls of the inner chamber. A temperature sensor (RTD PT100) was included in the fermentor that monitors the temperature of the inner chamber. The feature of the fermentor consisted of a heater and a pump connected to the outer chamber, circulating working fluid through the heater and the outer chamber and maintaining the temperatures of the inner chamber. In addition, a rotor blade was located within the inner chamber and connected to an electric motor; and a controller was configured to regulate the temperature within the inner chamber, the temperature of the working fluid, and the rotation speed of the electric motor.</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"186\" src=\"/media/article_images/2024/25/17/280-1722352436-Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure </strong><strong>1.</strong> Transparent CAD Model – the assemblage of the fermentor with bacterial growth chamber and rotary motion (Designed in SolidWorks V2021). A) Top view, B) Front view, and C) Dimetric view of the fermentor.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Description of the invention</strong></p>\r\n\r\n<p>The invention relates to the highly effective reliability of outputs with precision and a cost-efficient growth system with a significant decrease in production costs for biofertilizers (a consortium of bacterial cultures). The system was designed to produce approximately 25 liters of biofertilizer per production cycle. The heating system was embedded, and the temperature can be regulated, ranging from ambient temperature to 42°C. Furthermore, the speed of the rotor blade was adjusted through the RPM controller. The whole system operated from a 210-230V, 50-60Hz power source. This made the system user-friendly so that farmers could easily operate the setup under any load conditions in the rural areas of Bangladesh, where a limited supply of electricity is prevalent. Certain terminology was used to describe the instrument as depicted in <a href=\"#figure2\">Figure 2 </a>(A-E) for convenience only, and the numbering refers to the component of the instrument. In drawing, the terms ‘right,’ ‘left,’ ‘top,’ and ‘bottom' define the directional references. In addition, the words ‘a’ and ‘one' are noted as one or more of the items included unless specifically stated otherwise.</p>\r\n\r\n<p>The invention of a fermentor with a rotary motion and Heating system for the production of biofertilizers depicted in <a href=\"#figure2\">Figure 2</a> (A-E) contains (A) an outer chamber 1 made of aluminum, supporting the structure of the setup; (B) an inner bacterial growth chamber 3 made of aluminum – the working, chamber of the setup. The two chambers are welded together using an aluminum seal 2, keeping an equally distributed gap between the walls of the chambers. The temperature of the inner chamber is controlled by working fluid that flows between the gaps of the inner and outer chambers. Water is considered a working fluid that transfers heat from chamber 1 to chamber 3. The water was heated in heating chamber 7 (covered by a hood 6) and placed on fixed plate 5 of working chamber 3. The hot water was circulated between the two chambers by using a water pump 4. A set of pipes is connected between the water circulation area between chambers 1 and 3 through a 12-volt DC water pump 14 to the heating chamber 7. The water pump 14 was attached to the outer chamber 1 through a pump holder 15. The heating chamber 7 was fixed with the top fixed plate 5 through a chamber holder 8. A water heater 9 was placed inside the water heating chamber 7 and was regulated by a control panel 11 (AC 200-watt resistance heater) through a connecting wire 24. A rotor blade 13 was installed inside the growth chamber 3 and is connected to a 220V AC motor 12 operated by an AC power supply. This rotor blade 13 provides the rotary motion to the bacterial culture inside chamber 3. The rotor blade 13 is made of plastic so that it would not harm the bacteria growing inside chamber 3. A temperature sensor (RTD PT100) was installed inside chamber 3, recording the temperature of the culture medium, which needs to be controlled. The system was monitored by control unit 11 which harvests the signal from the temperature sensor installed inside chamber 3. The inner chamber 3 can be closed by a foldable hood 4 and the growth medium along with the bacterial inoculum can be inserted in the chamber by folding the hood. This foldable hood 4 was attached to the fixed hood 5 through a set of hinges 25. A set of fittings like flexible pipe 10, the pump inlet pipe 17, connecting pipe 18, tank fitting pipe 22, and different sets of joints 19, 20, and 21 were used for water circulation throughout the system. For power supply one main power cord 16 was connected to the control unit 11 which supplies power to the heater 9 and water pump 14 sequentially through connecting wires 23 and 24. Rotor blade 13 was powered through the main power cord 16, while a current regulator was added in the connection of the wire to the rotor blade 13 to control the speed of the rotor blade. All sets of connections were placed on the fixed plate 5. The temperature control unit of the system includes a temperature controller which was added to control unit 11, which has an external SSR (Solid State Relay) to realize a high heating power rate with optional PID (proportional–integral–derivative)<strong> </strong>control. Also, this device has an LCD (Liquid Crystal Display) with control switches for convenient operation. The controller has 250V AC 7.5A load for output and 20mA/10V DC Max load for SSR drive output. An NTC (Negative Temperature Coefficients), 5K/3470 sensor for temperature measurement was set. The operation range for temperature is 0°C -100°C with ± 1°C accuracy.</p>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"356\" src=\"/media/article_images/2024/25/17/280-1722352436-Figure2.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure </strong><strong>2</strong>. 2D drawing of the fermentor (Designed in SolidWorks 2021). A) Cross-sectional view that is consistent with the patent image [<a href=\"#r-33\">33</a>], B) Detailed sketch with an exploded view marking each component with a number, C) Water heating chamber with heater and lid, D) Rotor blade/ Agitator with motor, and E) Water circulating pump of the fermentor [<a href=\"#r-33\">33</a>].</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Working principle</strong></p>\r\n\r\n<p>To keep the temperature consistent, the system connects a temperature controller to the water heater and a sensor probe, taking the temperature reading directly from the bacterial growth chamber 3 (<a href=\"#figure2\">Figure 2</a>) while the water heater warms the water inside the heating chamber. The heating chamber, measuring 5 inches in length, 5 inches in width, and 12 inches in height, is filled with water. The water was heated by an induction water heater rated at 1500 watts. The heated water leaves the chamber through the outlet and enters the water jacket via a flexible pipe, causing the uniform heating of the bacterial culture in the chamber 3 (2). This heated water then circulates and transfers heat to the bacterial growth chamber until the desired temperature is reached. In addition, a propeller continuously rotates inside the chamber, preventing the bacterial bodies from conjugating simultaneously, ensuring the bacterial mixture is evenly heated, and allowing the bacteria to grow at a consistent rate.</p>\r\n\r\n<p><em> </em></p>\r\n\r\n<p><strong>Validation of temperature stability of the growth chamber</strong></p>\r\n\r\n<p>Three primary modes of energy transfer involve the movement of heat between two systems with different temperatures. The first mode was heating conduction, which refers to the transfer of heat through atomic vibration, typically in solid objects. The second mode was heating convection, which involves the transfer of heat through mass flow between systems and was a significant factor in heat transfer in liquid systems. The third mode was heat radiation, which involves the emission of electromagnetic radiation from any object with a temperature higher than absolute zero (-273.15℃/0 K) [<a href=\"#r-34\">34, 35</a>]<em>.</em> It was important to select the material used to build the chamber based on its thermal conductivity, calculated by determining the amount of heat that passes through a unit thickness of the material per unit area, given a specific temperature difference as previously reported [<a href=\"#r-36\">36</a>].</p>\r\n\r\n<p>We started the measurement of temperatures with our known temperature controller, but to check their accuracy, we used the Nicety DT1312 digital 2 input K-type thermometer. We recorded temperature values ranging from 28°C to 42°C, methodically regulating the agitator fan speeds in the chamber to 60, 75, and 90 rpm. Each temperature measurement was taken precisely to the closest 0.5°C, using the accuracy of the thermometer and a timer to track the time. We methodically recorded the temperature transitioning from 42°C to 28°C and noted every 0.5 increment change. This procedure allowed us to create a complete set of data, and to record with precision the intricate interplay between the temperature and fan speed within the chamber at any given time, achieving the desirable bacterial growth.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Nutrient substrates and bacterial consortia</strong></p>\r\n\r\n<p>The bacterial growth chamber was carefully assembled with the immobilizing structures and subjected to a thorough sterilization process to ensure the purity of the culture. The ingredients were mixed with 25 liters of water in a chamber and maintained at 37°C with agitation of 90 rpm, ensuring proper mixing and avoiding sedimentation. We have developed a new, affordable growth medium for large-scale cultures of next-generation bacterial biofertilizers. The medium consists of the ingredients with the amount as shown in <a href=\"#Table-1\">Table 1</a>.</p>\r\n\r\n<p>The following four endophytic bacterial consortia [<a href=\"#r-29\">29</a>] previously reported for rice plant growth were tested for their growth suitability in the newly assembled bacterial growth chamber for 72 h at 37ºC: Consortium A: <em>Klebsiella </em>sp. HSTU-Bk11 (MK656947.1), <em>Acinetobacter</em> sp. HSTU-Abk29 (MK695711.1), <em>Citrobacter</em> sp. HSTU-Abk30 (MK695712.1), and <em>Enterobacter cloacae</em> HSTU-Abk39 (MN795715); Consortium B: <em>Enterobacter cloacae</em> HSTU ABk37 (MN795712), <em>Enterobacter ludwigii</em> HSTU-Abk40 (MN795716), <em>Acinetobacter baumannii</em> HSTU-ABK42 (MN795722), <em>Klebsiella </em>sp. HSTU-ABk31 (MK695713.1), <em>Acinetobacter </em>sp. HSTU-Bk12 (MK656948.1); Consortium C: <em>Pseudomonas</em> sp. HSTU-Bk13 (MK656949.1), <em>Citrobacter</em> sp. HSTU-Bk14 (MK656950.1), <em>Acinetobacter </em>sp. HSTU-Bk15 (MK656951.1), <em>Acinetobacter </em>sp. HSTU-ABk32 (MN559060.1), and <em>Burkholderia </em>sp. HSTU-ABK33 (MK695714.1); and Consortium D: <em>Acinetobacter </em>sp. HSTU-ABk34 (MK695715.1), <em>Enterobacte</em>r sp. HSTU-ABk36 (MN795711), <em>Enterobacter </em>sp. HSTU-ABk38 (MN795713), and <em>Serratia marcescens</em> HSTU-ABk41 (795720). These strains were chosen for their compatibility in the consortium as previously reported [<a href=\"#r-29\">29</a>]. Each isolate was grown in nutrient broth for 24 h (10<sup>6</sup> CFU/mL for rice plants). We have used corn powder with commercial tryptic soya broth (TSB) and yeast extract (YE) powder in varied amounts (5-10 gm/25 liters) as reported previously [<a href=\"#r-29\">29</a>]. In addition, the growth of the tomato plant growth-promoting bacteria, <em>Enterobacter</em> sp. strain HSTU-ASh6 [<a href=\"#r-30\">30</a>] was conducted as described for the consortia of the endophytic bacteria for 72 h at 37ºC.</p>\r\n\r\n<p style=\"text-align:center\"><strong>Table </strong><strong>1</strong>. Culture medium ingredients.</p>\r\n\r\n<div id=\"Table-1\">\r\n<p style=\"text-align:center\"><a href=\"https://plant-trends.bsmiab.org/table/280-1722352436-table1/\"> Table-1</a></p>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Application of biofertilizer in plants</strong></p>\r\n\r\n<p>We have applied twice the amplified bacterial consortia-A to rice plants (var. BRRI dhan28) in fields where nitrogen fertilizer inputs were reduced by 60–80% of the recommended doses in order to demonstrate the effectiveness of the biogrowth chamber on the production of biofertilizer. The biofertilizer's ability to reduce nitrogen fertilizer was measured, as were the morphological growth parameters (plant heights, root length, shoot length, leaf breadth, and chlorophyll contents).</p>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p><strong>Thermal stability of growth chamber</strong></p>\r\n\r\n<p><a href=\"#figure3\">Figure 3</a> depicts the relationship between time and temperature for reaching the set value temperature (37°C) at three different agitation speeds of 60 rpm, or 75 rpm, or 90 rpm. The fastest gaining of preset temperature (37°C) was recorded to be at 90 rpm, followed by 75 rpm and then 60 rpm. A consistent trend of less than an hour to reach 37°C was recorded, indicating high efficiency and consistency of the system. The linear progression of time taken, and temperature rise implied a controlled and predictable heating process, demonstrating the stability and reliability of the system. This is critical for precise conditions required for the bacterial culture (production of bio-fertilizer), where temperature control is essential for optimal growth [<a href=\"#r-37\">37</a>].</p>\r\n\r\n<p>To examine the stability of temperature at the time of power disruption, a thorough examination of the cooling process was studied which reveals that the temperature of the medium drops from 37°C to 29°C approximately over four and a half hours, providing valuable insight into the underlying phenomenon. <a href=\"#figure4\">Figure 4</a> displays the cooling process dynamics within the system, illuminating an in-depth temperature-time relationship during the cooling phase. Unlike the heating process, the data indicated a gradual reduction in temperature from the initial 37°C to 29°C. This observed cooling pattern provides valuable insights into the efficiency and dynamics of the cooling mechanism.</p>\r\n\r\n<div id=\"figure3\">\r\n<figure class=\"image\"><img alt=\"\" height=\"401\" src=\"/media/article_images/2024/25/17/280-1722352436-Figure3.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure </strong><strong>3</strong>. Effect of RPM level on chamber temperature elevation. Temperature increases over time by the agitator/blade speed, operating at 60 RPM, 75 RPM, or 90 RPM to reach from 29°C to the required temperature of 37°C. Abbreviation, RPM, rotation per minute.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"figure4\">\r\n<figure class=\"image\"><img alt=\"\" height=\"404\" src=\"/media/article_images/2024/25/17/280-1722352436-Figure4.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure </strong><strong>4</strong>. Effect of RPM level on chamber temperature reduction. Temperature decreases over time by the agitator/blade speed, operating at 60 RPM, 75 RPM, or 90 RPM to cool down the temperature from 37°C to the initial temperature of 29°C. RPM, rotation per minute.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Streamline of working fluid</strong></p>\r\n\r\n<p>The Comsol Multiphysics v6.0 software was utilized to conduct a thorough simulation of the model under consideration. The simulation aimed to analyze the heated water flow around the growth chamber within the water jacket. The simulation results were presented through colored streamlines in <a href=\"#figure5\">Figure 5</a>. The illustration shows uneven water flow distribution around the jacketed system. Streamlines do not directly flow from the inlet to the outlet line, but circulate beneath the inlet, causing a complex flow pattern. Then the streamlines follow the side walls to the lower jacket and recirculate. The absence of streamlines navigating directly from the inlet to the outlet through the lower jacket indicates a distinct flow behavior. The streamlines exiting the system are mainly on the upper side, while the circulation streamlines are on the lower side, creating turbulence. Such type of flow ensures the good distribution of temperature in a growth chamber during water circulation [<a href=\"#r-38\">38</a>] which is crucial for maintaining an ideal environment within the chamber, ensuring all areas receive equal and regulated temperatures and producing an optimum growth of the bacterial culture.</p>\r\n\r\n<div id=\"figure5\">\r\n<figure class=\"image\"><img alt=\"\" height=\"359\" src=\"/media/article_images/2024/25/17/280-1722352436-Figure5.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure </strong><strong>5</strong>. The streamline of the circulating water around the inner chamber/growth chamber (Turbulent flow K-ɷ spf, Simulated in Comsol Multiphysics V6.0). A) Top view, B) Front view, C) Isometric view, and D) Side view of the fermentor.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>The growth rate of bacterial consortia</strong></p>\r\n\r\n<p><a href=\"#figure6\">Figure 6</a>A depicts the growth of all four consortia, A, B, C, and D in media containing both tryptic soya broth (5g) and yeast extract (5g). The OD values increased almost linearly from 0.25-0.5 to 1.75-2.2 throughout 12 to 48 hours of culture. From 48 to 60 hours, the growth rate significantly increased to 3 OD-3.5 OD. However, after 60 hours until 72 hours, the growth rate slows down and shows a horizontal trend with a growth rate (OD ranges 3.5~4.0). The <a href=\"#figure6\">Figure 6</a>B shows the growth of the consortia, A, B, C, and D in media containing only tryptic soya broth (5g). The OD values exhibited a steady increase from 0.25-0.5 to 0.75-1.25 during a period of 12 to 36 hours. Following 48 hours, the growth rate increased significantly to 1.75-2.0. However, after 60 hours, the growth rate accelerated rapidly, and the trend showed a vertical increase with a growth rate ranging from an optical density (OD) value 3.25 to 3.75. On the other hand, <a href=\"#figure6\">Figure 6</a>C demonstrates the growth of the consortia, A, B, C, and D in media containing only yeast extract (5g). The OD values showed a nearly linear increase over the entire growth period. The growth rate of consortia A, B, C, and D has been observed to increase OD value from 0.25-0.5 to 1.48-1.75 throughout the period of 12 to 48 hours. After 60 hours, the growth rate was observed OD value from 1.78 to 2.4. After 72 hours, the growth rate showed a range of OD value from 2.78 to 3.5. The consortia, A, B, C, and D in a medium containing no yeast extract and no tryptic soya broth showed a minimal growth ranging OD value from 1.75 - 2.25 (<a href=\"#figure6\">Figure 6</a>D).</p>\r\n\r\n<p>Using a combination of tryptic soya broth (5g) and yeast extract (5g) in a 25-litre sample, the growth rate of Enterobacter sp. strain HSTU-ASh6 was observed (<a href=\"#figure6\">Figure 6</a>E). The OD value increased from 0.25 to 3.75 throughout 12 to 72 hours. The growth rate was found to be highly accelerated after 36 hours.</p>\r\n\r\n<p>Overall, consortia of endophytic bacteria grown in media containing TSB and YE exhibited robust growth over the incubation period. In contrast, when either TSB or YE was absent from the medium a noticeable reduction in growth rate was observed for all consortia. The absence of both TSB and YE led to a marked decline in growth across all consortia, indicating the essential role of these components in bacterial proliferation. Further optimization experiments revealed that while both TSB and YE contributed to optimal growth, YE supplementation can be minimized without significantly compromising growth rates. Consortia grown without YE exhibited slightly reduced growth rates compared to those grown with both TSB and YE, but the difference was not substantial, suggesting that YE supplementation can be omitted to reduce production costs while still maintaining the satisfactory growth performance of the consortia. Interestingly, when a tomato plant growth-promoting endophytic bacteria, <em>Enterobacter</em> sp. strain HSTU-ASh6 was cultured in the bacterial growth chamber for 72 hours in presence of both TSB and YE, the high growth rate of the strain (<a href=\"#figure6\">Figure 6</a>E) was noted like other consortia A, B, C and D (<a href=\"#figure6\">Figure 6</a>A-D).</p>\r\n\r\n<div id=\"figure6\">\r\n<figure class=\"image\"><img alt=\"\" height=\"523\" src=\"/media/article_images/2024/25/17/280-1722352436-Figure6.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure </strong><strong>6</strong>. The growth rate of consortia A, B, C, and D. A) Growth medium containing tryptic soya broth (TSB=5.0 g) and yeast extract (5 g), OD = 10 g, volume = 25 liters; B) Growth medium containing only tryptic soya broth (TSB= 5.0 g), OD = 10 g, volume = 25 liters; C) Growth medium containing only yeast extract, OD = 5.0 g, volume = 25 liters; D) Growth medium without consisting of TSB and Yeast extract. OD = 10 g, volume = 25 liters; E) Growth rate of Enterobacter sp. strain HSTU-ASh6 in medium containing tryptic soya broth (TSB=5.0 g) and yeast extract (5 g), OD = 10 g, volume = 25 liters.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Plant growth promotion as a biofertilizer</strong></p>\r\n\r\n<p>As shown in <a href=\"#Table-1\">Table 2</a> and <a href=\"#figure7\">Figure 7</a>, the rice plants' height, root length, and number of tillers were all improved by the bacterial consortia-A treatment. Notably, compared to all other treatments, the number of tillers at 70% urea reduction (Treatment B) linked to the application of biofertilizer consortia A was noticeably higher (<a href=\"#Table-2\">Table 2</a>). In a similar vein, the rice plants in treatment A had the longest roots. On the other hand, treatment E was determined to have the tallest plants. These findings suggested that even in the reduction of urea fertilizer, the biofertilizer potential bacterial consortia A considerably accelerated the morphological features of rice plants associated with yields.</p>\r\n\r\n<p style=\"text-align:center\"><strong>Table 2.</strong> Rice plants (var. BRRI dhan28) morphological traits in response to the biofertilizer consortia amplified in the newly developed fermentor.</p>\r\n\r\n<div id=\"Table-2\">\r\n<p style=\"text-align:center\"><a href=\"https://plant-trends.bsmiab.org/table/280-1722352436-table2/\">Table-2</a></p>\r\n</div>\r\n\r\n<div id=\"figure7\">\r\n<figure class=\"image\"><img alt=\"\" height=\"414\" src=\"/media/article_images/2024/25/17/280-1722352436-Figure7.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 7</strong>. Growth performance of amplified biofertilizer consortia A on rice plants (var. BRRI dhan28) in field conditions with reduced urea fertilizer. The morphology of rice plants was attained with A) 40%, B) 30%, C) 20%, D) 0%, and E) 100% of the recommended dose of urea, respectively, where bacterial consortia A were applied twice in the treatments A, B, and C.</figcaption>\r\n</figure>\r\n</div>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p>The invention of an efficient reliable fermentor with growth chambers equipped with heating and rotary motion pertains to enhanced biofertilizer production (consortia of endophytic bacteria). Biofertilizer growth chambers are devices used in the fields of biochemistry, molecular biology, bacteriology, and organic farming [<a href=\"#r-38\">38</a>]. Continuous motion not only prevents precipitation but also ensures the circulation of solvents over all the regions of test samples, which ensures better mixing [<a href=\"#r-40\">40</a>]. Production of bacteria in large quantities can be difficult and expensive, due to the size and cost of currently available growth systems including the culture media, as well as the availability of such systems primarily in first-world countries and the need to transport such currently available systems to other regions of the world. Therefore, a need exists for a fermentor for bio-fertilizer production that can provide heating and rotary motion in controlled conditions while remaining cost-effective and portable. Thus, to meet this challenge an affordable fermentor is well desired.</p>\r\n\r\n<p>By monitoring the temperature directly from the bacterial growth chamber and heating the water in the heating chamber using an induction water heater, we can ensure that the temperature remains consistent throughout the growth process. However, slight variations have been seen due to the differences in the agitation (rpm) of the propeller. The variations in the rpm have a direct impact on the convection on the surface of the culture, leading to slight temperature variations [<a href=\"#r-41\">41</a>]. Again, this finding is of utmost importance as it underscores the fact that the culture medium within the growth chamber can maintain its stability even in the event of a brief power disruption as the cooling process takes a very long time. This serves to highlight the prototype’s resilience and its capacity to endure minor disturbances with minimal impact. In addition, the capacity to distribute temperature uniformly is highly desirable for uniform bacterial growth. Indeed, we demonstrated that the prototype is reliable, and functioning efficiently. Several devices like ours use water jacket heating systems for a specific purpose, including the fermentation tank improvement structure, convenient for fixed stirring devices [<a href=\"#r-42\">42</a>]; a dry powder composite coating prepared [<a href=\"#r-43\">43</a>]; and pasteurizing apparatus [<a href=\"#r-44\">44</a>].</p>\r\n\r\n<p>A similar concept of fermentor illustrated by Yang [<a href=\"#r-45\">45</a>] uses a water jacket for uniform heat distribution to the inner chamber. However, it does not include any integrated heating source, nor does it include a pump that is fluidly connected to the outer chamber to circulate the working fluid between the heater and the outer chamber. If we want to control the temperature of the growth chamber proposed by Yang, an externally heat-controlled water flow must be introduced. Furthermore, Yang’s model includes a complex set of mixing rods, which makes the system more difficult to manufacture, increasing the manufacturing cost and making it unaffordable for the farmers. On the other hand, the agitator used in our concept is designed in such a way that it can mix the bacterial consortium easily, an essential property deals with different types of husks of the members of the consortium, that which makes the liquid thicker. Our prototype fermentor uses a simple single motor-controlled propeller that makes it easier to build. While Yang’s Model showed the inlet and outlet of the working fluid are placed on the same side which may not result in a good fluid flow around the growth chamber, hindering the bacterial growth, the prototype presented here has the inlet and outlet on the opposite side which showed a good flow of the working fluid around the growth chamber as the simulated contour shows (<a href=\"#figure5\">Figure 5</a>). Furthermore, our prototype demonstrates how effectively plant growth-promoting bacteria can be grown to accelerate agricultural productivity.</p>\r\n\r\n<p>The system can be further modified to its range of production and temperature control. Recently, some other cost-effective concepts have been deployed to make fermentors for large-scale biofertilizer production [<a href=\"#r-46\">46</a>] with a limit of 3.8 L biofertilizer per cycle, whereas our prototype has a capacity of 25 L per cycle with thermal stability under a power disruption. These characteristics of our compact and unique fermentor provide a design for the first-ever reported. Hence, we have patented the design under the United States Patent and Trademark Office (USPTO) [<a href=\"#r-33\">33</a>]. The results of our study emphasized the critical role of tryptic soya broth (TSB) and yeast extract (YE) in promoting the growth of endophytic bacterial consortium in the prototype fermentor. Both single endophytic strains and consortia of different strains can adopt the growth media in the growth chamber without losing their properties. The observed robust growth of consortia in media containing both TSB and YE highlights the synergistic effects of these components on bacterial proliferation. TSB provides essential nutrients, including peptides, amino acids, and carbohydrates, while YE supplies vitamins, minerals, and growth factors necessary for bacterial growth. The combination of these components creates an optimal milieu for bacterial growth, enabling consortia to thrive and proliferate as observed [<a href=\"#r-29\">29, 30</a>]. Furthermore, our optimization experiments revealed that YE supplementation can be minimized without significantly compromising growth rates. Consortia grown without YE exhibited slightly reduced growth rates compared to those grown with both TSB and YE, indicating that YE supplementation may be partially expendable for cost-saving purposes. This finding is particularly relevant for large-scale production efforts of biofertilizers, where reducing production costs is a priority. In the future, it is anticipated that research efforts will be directed toward refining media formulations to achieve optimal growth performance. This will involve exploring alternative nutrient sources to further enhance growth and simultaneously reduce production costs, helping sustainable organic farming.</p>\r\n\r\n<p>The present study revealed a cheap formulation of growth medium can be utilized for the amplification of biofertilizer production using the newly developed prototype fermentor. This invention helps local farmers or industries to set up a sustainable technology for large-scale biofertilizer production cheaply, enhancing the crop’s yields.</p>\r\n\r\n<p>The consortia (A-D) were reported to increase the rice (BRRI-dhan28) production in field conditions by 28-35% [<a href=\"#r-29\">29</a>]. The production of tomatoes in field conditions increased 2.5-fold when treated with the single endophytic strain <em>Enterobacter</em> sp. HSTU-ASh6 [<a href=\"#r-30\">30</a>]. Both rice and tomato production were increased in the presence of a 70% reduction of the recommended doses of urea and no pesticides [<a href=\"#r-29\">29</a>]. Therefore, amplification of the bacterial strains (biofertilizers) in the novel fermentor with a cheap growth medium provided opportunities for developing sustainable organic agricultural practices. Using this technology, we have conducted several crop and vegetable field trials with various sizes of endophytic consortia that can replace numerous amounts of agrochemicals, e.g., phosphate, urea, pesticides, and increase biofunctional compounds (data unpublished). Therefore, the study provides new insights for researchers and industrialists regarding biofertilizers to ensure the sustainable development of organic agriculture and improve farming by producing hazard-free crops. The consortia A amplified in the biogrowth chamber showed noticeable improvements in the rice plant’s morphological traits (<a href=\"#Table-1\">Table 1</a> and <a href=\"#figure7\">Figure 7</a>) related to yields, which indicated that the biogrowth chamber created an optimal environment for the production as an endophytic bacterial-based biofertilizer.</p>"
},
{
"section_number": 5,
"section_title": "CONCLUSIONS",
"body": "<p>This study explored a newly designed fermentor (bacterial growth chamber) as well as the newly discovered affordable medium for the production of biofertilizers for environmentally friendly organic farming. The growth medium is significantly cheaper than traditional media used for large-scale cultures of bacteria. Furthermore, the prototype fermentor has been demonstrated to be sustainable, as it uses waste products from the food industry, such as poultry feed and corn flour. We have successfully demonstrated that this new medium is used to culture a variety of next-generation plant growth-promoting bacteria in the consortium that can be used as biofertilizers for sustainable farming. This new, affordable growth medium has the potential to revolutionize the production of bacterial biofertilizers, making it more affordable and accessible to farmers around the world. The bacterial growth chamber reported in this paper is designed to provide an improved environment to produce biofertilizers. One of the main advantages of this fermentor is its ability to maintain a stable thermal environment during a prolonged power disruption, which is important for steady bacterial growth. The chamber is also designed to provide improved sterile conditions, reducing the risk of contamination and ensuring the purity of the final product. Another key feature of this prototype fermentor is its seamless operation, ensuring the growth process can proceed without interruption, allowing for efficient and consistent production of biofertilizers. Furthermore, the chamber is simple and easy to fabricate, with a low cost of approximately US $250, making it accessible and affordable for a wide range of users, including small-scale farmers. In particular, the rice plants morphological traits improvement with bacterial consortia application might be a milestone in future research for farmers practice for sustainable crop cultivation. </p>"
},
{
"section_number": 6,
"section_title": "ACKNOWLEDGMENTS",
"body": "<p>This research was partially funded by grants from the Institute of Research and Training (IRT)-Hajee Mohammad Danesh Science and Technology University, Dinajpur, Bangladesh. The fermentor invention was patented by the United States Patent and Trademark Office (title ‘Biogrowth chamber with rotary motion and heating system and related methods’ Application number: 18/409771, priority date: January 10, 2024), Patent number US12,084,644B1 (<a href=\"https://patentcenter.uspto.gov/applications/18409771\">https://patentcenter.uspto.gov/applications/18409771</a>).</p>"
},
{
"section_number": 7,
"section_title": "AUTHOR CONTRIBUTIONS",
"body": "<p>TA: Invention, Methodology, Investigation, Analysis, Writing; MSR: Invention, Methodology, Investigation, Analysis, Design and Simulation, Writing and Editing; R: Methodology, Writing; SG: Consulting, Review and Editing; MMH: Data Analysis; MAH: Invention, Conceptualization, Supervision, Project administration, Writing, Review and Editing; AR: Invention, Review and Editing. All authors have approved the final version.</p>"
},
{
"section_number": 8,
"section_title": "CONFLICTS OF INTEREST",
"body": "<p>There is no conflict of interest among the authors.</p>"
}
],
"figures": [
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/25/17/280-1722352436-Figure1.jpg",
"caption": "Figure 1. Transparent CAD Model – the assemblage of the fermentor with bacterial growth chamber and rotary motion (Designed in SolidWorks V2021). A) Top view, B) Front view, and C) Dimetric view of the fermentor.",
"featured": false
},
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/25/17/280-1722352436-Figure2.jpg",
"caption": "Figure 2. 2D drawing of the fermentor (Designed in SolidWorks 2021). A) Cross-sectional view that is consistent with the patent image [33], B) Detailed sketch with an exploded view marking each component with a number, C) Water heating chamber with heater and lid, D) Rotor blade/ Agitator with motor, and E) Water circulating pump of the fermentor [33].",
"featured": false
},
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/25/17/280-1722352436-Figure3.jpg",
"caption": "Figure 3. Effect of RPM level on chamber temperature elevation. Temperature increases over time by the agitator/blade speed, operating at 60 RPM, 75 RPM, or 90 RPM to reach from 29°C to the required temperature of 37°C. Abbreviation, RPM, rotation per minute.",
"featured": false
},
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/25/17/280-1722352436-Figure4.jpg",
"caption": "Figure 4. Effect of RPM level on chamber temperature reduction. Temperature decreases over time by the agitator/blade speed, operating at 60 RPM, 75 RPM, or 90 RPM to cool down the temperature from 37°C to the initial temperature of 29°C. RPM, rotation per minute.",
"featured": false
},
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/25/17/280-1722352436-Figure5.jpg",
"caption": "Figure 5. The streamline of the circulating water around the inner chamber/growth chamber (Turbulent flow K-ɷ spf, Simulated in Comsol Multiphysics V6.0). A) Top view, B) Front view, C) Isometric view, and D) Side view of the fermentor.",
"featured": false
},
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/25/17/280-1722352436-Figure6.jpg",
"caption": "Figure 6. The growth rate of consortia A, B, C, and D. A) Growth medium containing tryptic soya broth (TSB=5.0 g) and yeast extract (5 g), OD = 10 g, volume = 25 liters; B) Growth medium containing only tryptic soya broth (TSB= 5.0 g), OD = 10 g, volume = 25 liters; C) Growth medium containing only yeast extract, OD = 5.0 g, volume = 25 liters; D) Growth medium without consisting of TSB and Yeast extract. OD = 10 g, volume = 25 liters; E) Growth rate of Enterobacter sp. strain HSTU-ASh6 in medium containing tryptic soya broth (TSB=5.0 g) and yeast extract (5 g), OD = 10 g, volume = 25 liters.",
"featured": false
},
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/25/17/280-1722352436-Figure7.jpg",
"caption": "Figure 7. Growth performance of amplified biofertilizer consortia A on rice plants (var. BRRI dhan28) in field conditions with reduced urea fertilizer. The morphology of rice plants was attained with A) 40%, B) 30%, C) 20%, D) 0%, and E) 100% of the recommended dose of urea, respectively, where bacterial consortia A were applied twice in the treatments A, B, and C.",
"featured": true
}
],
"authors": [
{
"id": 68,
"affiliation": [
{
"affiliation": "Department of Biochemistry and Molecular Biology, Faculty of Agriculture, Hajee Mohammad Danesh Science and Technology University, Dinajpur 5200, Bangladesh"
},
{
"affiliation": "Department of Mechanical Engineering, Faculty of Engineering, Hajee Mohammad Danesh Science and Technology University, Dinajpur 5200, Bangladesh"
}
],
"first_name": "Tanweer",
"family_name": "Ahmed",
"email": null,
"author_order": 1,
"ORCID": null,
"co_first_author": true,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 15
},
{
"id": 69,
"affiliation": [
{
"affiliation": "Department of Mechanical Engineering, Faculty of Engineering, Hajee Mohammad Danesh Science and Technology University, Dinajpur 5200, Bangladesh"
},
{
"affiliation": "Department of Biochemistry and Molecular Biology, Faculty of Agriculture, Hajee Mohammad Danesh Science and Technology University, Dinajpur 5200, Bangladesh"
}
],
"first_name": "Md Shahriar",
"family_name": "Rahman",
"email": null,
"author_order": 2,
"ORCID": null,
"co_first_author": true,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 15
},
{
"id": 71,
"affiliation": [
{
"affiliation": "Department of Mechanical Engineering, Faculty of Engineering, Hajee Mohammad Danesh Science and Technology University, Dinajpur 5200, Bangladesh"
}
],
"first_name": "Rokanuzzaman",
"family_name": null,
"email": null,
"author_order": 3,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 15
},
{
"id": 72,
"affiliation": [
{
"affiliation": "Albany College of Pharmacy and Health Sciences, 106 New Scotland Avenue, Albany, NY 12208, United States of America"
}
],
"first_name": "Sibdas",
"family_name": "Ghosh",
"email": null,
"author_order": 4,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 15
},
{
"id": 73,
"affiliation": [
{
"affiliation": "Department of Biomedical Sciences, King Faisal University, Al-Hofuf, Al-Ahsa, Saudi Arabia"
}
],
"first_name": "Aminur",
"family_name": "Rahman",
"email": null,
"author_order": 5,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 15
},
{
"id": 74,
"affiliation": [
{
"affiliation": "Department of Mechanical Engineering, Faculty of Engineering, Hajee Mohammad Danesh Science and Technology University, Dinajpur 5200, Bangladesh"
}
],
"first_name": "Md. Mehdi",
"family_name": "Hasan",
"email": null,
"author_order": 6,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 15
},
{
"id": 70,
"affiliation": [
{
"affiliation": "Department of Biochemistry and Molecular Biology, Faculty of Agriculture, Hajee Mohammad Danesh Science and Technology University, Dinajpur 5200, Bangladesh"
}
],
"first_name": "Md. Azizul",
"family_name": "Haque",
"email": "helalbmb2016@hstu.ac.bd",
"author_order": 7,
"ORCID": "https://orcid.org/0000-0002-9788-0766",
"co_first_author": false,
"co_author": false,
"corresponding": true,
"corresponding_author_info": "Md. Azizul Haque,PhD; Department of Biochemistry and Molecular Biology, Faculty of Agriculture, Hajee Mohammad Danesh Science and Technology University, Dinajpur 5200, Bangladesh. Email: helalbmb2016@hstu.ac.bd",
"article": 15
}
],
"views": 1045,
"downloads": 249,
"references": [
{
"id": 518,
"serial_number": 1,
"reference": "Duro JA, Lauk C, et al. Global inequalities in food consumption, cropland demand and land-use efficiency: A decomposition analysis. Global Environmental Change. 2020;64:102124.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 519,
"serial_number": 2,
"reference": "UN. United nations world population report. 2023.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 520,
"serial_number": 3,
"reference": "van Dijk M, Morley T, et al. A meta-analysis of projected global food demand and population at risk of hunger for the period 2010–2050. Nature Food. 2021;2:494-501.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 521,
"serial_number": 4,
"reference": "Fukase E, Martin W. Economic growth, convergence, and world food demand and supply. World Development. 2020;132:104954.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 522,
"serial_number": 5,
"reference": "Pahalvi HN, Rafiya L, et al. Chemical fertilizers and their impact on soil health. Microbiota and Biofertilizers, Vol 2: Ecofriendly tools for reclamation of degraded soil environs. 2021:1-20.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 523,
"serial_number": 6,
"reference": "Rashmi I, Roy T, et al. Organic and inorganic fertilizer contaminants in agriculture: Impact on soil and water resources. Contaminants in Agriculture: Sources, Impacts and Management. 2020:3-41.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 524,
"serial_number": 7,
"reference": "Planning Mo. Foreign trade stastics of bangladesh. 2022-23.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 525,
"serial_number": 8,
"reference": "Walker DJ, Clemente R, et al. Contrasting effects of manure and compost on soil ph, heavy metal availability and growth of chenopodium album l. In a soil contaminated by pyritic mine waste. Chemosphere. 2004;57:215-24.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 526,
"serial_number": 9,
"reference": "Amoo AE, Enagbonma BJ, et al. Biofertilizer: An eco-friendly approach for sustainable crop production. Food Security and Safety: African Perspectives. 2021:647-69.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 527,
"serial_number": 10,
"reference": "Verma DK, Pandey AK, et al. Plant growth-promoting rhizobacteria: An eco-friendly approach for sustainable agriculture and improved crop production. Microbiology for sustainable agriculture, soil health, and environmental protection: Apple Academic Press; 2019. p. 3-80.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 528,
"serial_number": 11,
"reference": "Ha-Tran DM, Nguyen TTM, et al. Roles of plant growth-promoting rhizobacteria (pgpr) in stimulating salinity stress defense in plants: A review. International Journal of Molecular Sciences. 2021;22:3154.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 529,
"serial_number": 12,
"reference": "Enebe MC, Babalola OO. The influence of plant growth-promoting rhizobacteria in plant tolerance to abiotic stress: A survival strategy. Applied microbiology and biotechnology. 2018;102:7821-35.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 530,
"serial_number": 13,
"reference": "Etesami H, Adl SM. Plant growth-promoting rhizobacteria (pgpr) and their action mechanisms in availability of nutrients to plants. Phyto-Microbiome in stress regulation. 2020:147-203.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 531,
"serial_number": 14,
"reference": "Nagrale D, Chaurasia A, et al. Pgpr: The treasure of multifarious beneficial microorganisms for nutrient mobilization, pest biocontrol and plant growth promotion in field crops. World Journal of Microbiology and Biotechnology. 2023;39:100.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 532,
"serial_number": 15,
"reference": "Di Benedetto NA, Corbo MR, et al. The role of plant growth promoting bacteria in improving nitrogen use efficiency for sustainable crop production: A focus on wheat. AIMS microbiology. 2017;3:413.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 533,
"serial_number": 16,
"reference": "Verma M, Mishra J, et al. Plant growth-promoting rhizobacteria: Diversity and applications. Environmental biotechnology: for sustainable future. 2019:129-73.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 534,
"serial_number": 17,
"reference": "Chauhan H, Bagyaraj D, et al. Novel plant growth promoting rhizobacteria—prospects and potential. Applied Soil Ecology. 2015;95:38-53.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 535,
"serial_number": 18,
"reference": "Khan N, Bano A, et al. Crosstalk amongst phytohormones from planta and pgpr under biotic and abiotic stresses. Plant Growth Regulation. 2020;90:189-203.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 536,
"serial_number": 19,
"reference": "Bhat M, Rasool R, et al. Plant growth promoting rhizobacteria (pgpr) for sustainable and eco-friendly agriculture. Acta Sci Agric. 2019;3:23-5.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 537,
"serial_number": 20,
"reference": "Martínez-Viveros O, Jorquera MA, et al. Mechanisms and practical considerations involved in plant growth promotion by rhizobacteria. Journal of soil science and plant nutrition. 2010;10:293-319.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 538,
"serial_number": 21,
"reference": "Etesami H. Plant–microbe interactions in plants and stress tolerance. Plant life under changing environment: Elsevier; 2020. p. 355-96.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 539,
"serial_number": 22,
"reference": "Asghari B, Khademian R, et al. Plant growth promoting rhizobacteria (pgpr) confer drought resistance and stimulate biosynthesis of secondary metabolites in pennyroyal (mentha pulegium l.) under water shortage condition. Scientia Horticulturae. 2020;263:109132.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 540,
"serial_number": 23,
"reference": "Abhilash P, Dubey RK, et al. Plant growth-promoting microorganisms for environmental sustainability. Trends in Biotechnology. 2016;34:847-50.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 541,
"serial_number": 24,
"reference": "Ojkic N, Lilja E, et al. A roadblock-and-kill model explains the dynamical response to the DNA-targeting antibiotic ciprofloxacin. BioRxiv. 2019:791145.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 542,
"serial_number": 25,
"reference": "Maier RM, Pepper IL. Bacterial growth. Environmental microbiology: Elsevier; 2015. p. 37-56.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 543,
"serial_number": 26,
"reference": "Pörtner R, Nagel-Heyer S, et al. Bioreactor design for tissue engineering. Journal of bioscience and bioengineering. 2005;100:235-45.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 544,
"serial_number": 27,
"reference": "Lara AR, Galindo E, et al. Living with heterogeneities in bioreactors: Understanding the effects of environmental gradients on cells. Mol Biotechnol. 2006;34:355-81.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 545,
"serial_number": 28,
"reference": "Vassilev N, Vassileva M, et al. Unexploited potential of some biotechnological techniques for biofertilizer production and formulation. Applied microbiology and biotechnology. 2015;99:4983-96.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 546,
"serial_number": 29,
"reference": "Prodhan MY, Rahman MB, et al. Characterization of growth-promoting activities of consortia of chlorpyrifos mineralizing endophytic bacteria naturally harboring in rice plants—a potential bio-stimulant to develop a safe and sustainable agriculture. Microorganisms. 2023;11:1821.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 547,
"serial_number": 30,
"reference": "Haque MA, Hossain MS, et al. Unveiling chlorpyrifos mineralizing and tomato plant-growth activities of enterobacter sp. Strain hstu-ash6 using biochemical tests, field experiments, genomics, and in silico analyses. Frontiers in Microbiology. 2022;13:1060554.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 548,
"serial_number": 31,
"reference": "Baddoo N. Stainless steel in construction: A review of research, applications, challenges and opportunities. Journal of constructional steel research. 2008;64:1199-206.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 549,
"serial_number": 32,
"reference": "Tran T, Kannoorpatti K, et al. A study of bacteria adhesion and microbial corrosion on different stainless steels in environment containing desulfovibrio vulgaris. Royal Society open science. 2021;8:201577.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 550,
"serial_number": 33,
"reference": "Haque MA. Biogrowth chamber with rotary motion and heating system and related methods’ US patent 2024, Patent number US12,084,644B1 (https://patentcenter.uspto.gov/applications/18409771)",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 551,
"serial_number": 34,
"reference": "Janna WS. Engineering heat transfer: CRC press; 2018.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 552,
"serial_number": 35,
"reference": "Sidebotham G. Heat transfer modes: Conduction, convection, and radiation. In: Sidebotham G, editor. Heat transfer modeling: An inductive approach. Cham: Springer International Publishing; 2015. p. 61-93.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 553,
"serial_number": 36,
"reference": "Cengel Y, Cimbala J, et al. Ebook: Fundamentals of thermal-fluid sciences (si units): McGraw Hill; 2012.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 554,
"serial_number": 37,
"reference": "Moon S, Ham S, et al. Temperature matters: Bacterial response to temperature change. Journal of Microbiology. 2023;61:343-57.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 555,
"serial_number": 38,
"reference": "Patil PM, Yadav AP, et al. Comparative study between heat transfer through laminar flow and turbulent flow. Int J Innov Res Sci Eng Technol. 2015;4:2223-6.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 556,
"serial_number": 39,
"reference": "Mahanty T, Bhattacharjee S, et al. Biofertilizers: A potential approach for sustainable agriculture development. Environmental Science and Pollution Research. 2017;24:3315-35.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 557,
"serial_number": 40,
"reference": "Yunus MU, Silas K, et al. A review of biofertilizer production: Bioreactor, feedstocks and kinetics. 2022.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 558,
"serial_number": 41,
"reference": "Agrawal S, Simon T, et al. An experimental study on the effects of agitation on convective heat transfer. International Journal of Heat and Mass Transfer. 2015;90:302-13.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 559,
"serial_number": 42,
"reference": "Liao C-b. Fermentation tank improvement structure convenient for fixd stirring device China2016.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 560,
"serial_number": 43,
"reference": "Li W-j. A dry powder composite coating prepared by the preparation method and its device. China2020.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 561,
"serial_number": 44,
"reference": "J.W. Ladd HBW. Pasteurizing apparatus. USA1932.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 562,
"serial_number": 45,
"reference": "Yang Z, Qu. A multi-bacteria fusion symbiotic fermentation technique for disease resistance and promoting growth. China2021.",
"pmc": null,
"DOI": null,
"article": 15
},
{
"id": 563,
"serial_number": 46,
"reference": "Manzano-Gómez LA, Rincón-Rosales R, et al. Cost-effective cultivation of native pgpb sinorhizobium strains in a homemade bioreactor for enhanced plant growth. Bioengineering. 2023;10:960.",
"pmc": null,
"DOI": null,
"article": 15
}
]
},
{
"id": 13,
"slug": "280-1711854419",
"featured": true,
"slider": true,
"issue": "Vol2 Issue3",
"type": "review_article",
"manuscript_id": "280-1711854419",
"recieved": "2024-03-31",
"revised": null,
"accepted": "2024-06-24",
"published": "2024-09-25",
"pdf_file": "https://plant-trends.bsmiab.org/media/pdf_file/2024/30/280-1711854419.pdf",
"title": "Plant improvement and metabolite production in <span>Cannabis sativa</span>: Recent biotechnological advances",
"abstract": "<p>The <em>Cannabis sativa </em>plant is an excellent source of metabolites, fiber, and medicinal properties. Phytocannabinoids are the secondary metabolites naturally derived from <em>Cannabis</em> plant species<em>. </em>These metabolites are promising and can be used in producing phytomedicine or plant-based therapeutics. However, many of these compounds are produced in low quantities across different<em> Cannabis</em> species. To solve this limitation, <em>in vitro,</em> biotechnological methods offer promising solutions for enhancing the production of secondary metabolites in <em>Cannabis</em>. This review highlights the biotechnological approaches for enhancing <em>Cannabis </em>secondary metabolite production through <em>in vitro </em>plant improvement techniques such as plant regeneration, elicitor-responsive metabolite induction, polyploidy manipulation, protoplast culture, bioreactor-based hairy root culture, genetic transformation, and genome editing. These biotechnological approaches might be useful for improving <em>Cannabis</em> plants and increasing plant capacity to produce potential metabolites. These phytochemical and bioactive compounds found in <em>Cannabis</em> species could be used as alternative resources for pharmaceutical and industrial production.</p>",
"journal_reference": "Plant Trends. 2024; 2(3): 74-91",
"academic_editor": "Monirul Islam, PhD; University of Massachusetts Amherst Amherst, USA",
"cite_info": "Plant improvement and metabolite production in Cannabis sativa: Recent biotechnological advances. Plant Trends. 2024 Sep; 2(3): 74-91.",
"keywords": [
"Plant improvement",
"Elicitor",
"Secondary metabolite",
"Cannabis",
"Gene editing"
],
"DOI": "10.5455/pt.2024.07",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p><em>Cannabis sativa </em>L.<em> </em>is a high-value multipurpose plant. In early societies, this plant was used for medicinal benefits, and recreational elements, as a source of food additives, cosmetics, paper, bioenergy, and textiles [<a href=\"#r-1\">1</a>]. Additionally, <em>Cannabis</em> possesses pharmacological benefits, such as anti-cancer, anti-inflammatory, antispastic, anticonvulsant, anti-pruritic, and anti-psychotic elements [<a href=\"#r-2\">2</a>]. It is also used for ornamental and landscaping purposes [<a href=\"#r-3\">3</a>]. Phytocannabinoids are the primary secondary metabolites found in <em>Cannabis</em>, and are primarily formed in the glandular trichomes of female <em>Cannabis</em> flowers [<a href=\"#r-4\">4</a>]. Over 150 phytocannabinoids have been identified in <em>Cannabis</em> plants [<a href=\"#r-5\">5</a>]. Research associated with signalling pathways of phytocannabinoids leads to the identification of cannabinoid receptors and their endogenous ligands [<a href=\"#r-6\">6-9</a>]. These receptors have been found in various cellular compartments, which are associated with brain disorders, metabolic pathways, and immune function [<a href=\"#r-8\">8</a>]. Some cannabinoids are used for potential therapeutic use in treating COVID-19 [<a href=\"#r-10\">10-12</a>]. The plant containing cannabinoids, terpenes, and phenolic compounds is increased due to its pharmacological applications [<a href=\"#r-13\">13, 14</a>]. Breeding of <em>Cannabis</em> cultivars with specific secondary metabolite profiles for medicinal purposes is a slow process due to dioecy and regulatory constraints [<a href=\"#r-15\">15, 16</a>]. Therefore, alternative strategies for the rapid and efficient production of minor cannabinoids (CBG, CBN, CBC, and THCV), which are produced in lower quantities than major cannabinoids (THCA, CBDA), are of particular research interest [<a href=\"#r-17\">17</a>].</p>\r\n\r\n<p><em>In vitro</em> tissue culture methods, including callus, cell culture, <em>de novo</em> regeneration, hairy root culture, and protoplast culture, are essential for micropropagation [<a href=\"#r-18\">18</a>], and genetic engineering techniques (e.g., <em>Agrobacterium</em>-mediated gene transformation, <em>A. rhizogenes</em>-mediated hairy root cultures, etc.) as well as polyploidy induction can be used to improve traits with producing secondary metabolites in plants [<a href=\"#r-19\">19</a>]. These traditional methods can alter secondary metabolite production, the CRISPR/Cas9 system is one of the most significant approaches for faster genetic manipulation in <em>Cannabis</em>, especially in overcoming <em>in vitro</em> recalcitrance [<a href=\"#r-20\">20, 21</a>]. In this updated study, we focused on advances of <em>Cannabis</em> biotechnology approaches related to plant improvement and potential metabolite production, which will open a new era of <em>Cannabis </em>research.</p>"
},
{
"section_number": 2,
"section_title": "BIOSYNTHETIC PATHWAYS FOR METABOLITES PRODUCTION",
"body": "<p>Two main pathways are involved in the synthesis of cannabinoids in <em>Cannabis </em>plants. The polyketide pathway that produces olivetolic acid (OLA), and the plastidal 2-C-methyl-d-erythritol 4-phosphate (MEP) pathway that synthesises geranyl diphosphate (GPP). OLA originates from hexanoyl-CoA through aldol condensation with three molecules of malonyl-CoA by a polyketide synthase (PKS) enzyme and an olivetolic acid cyclase (OAC) through polyketide pathway (PKP) (<a href=\"#figure1\">Figure 1</a>). The alkylation of OLA with GPP by geranyl pyrophosphate: olivetolate geranyl transferase forms the central precursor CBGA. Oxidocyclases such as THCA synthase (THCAS), CBDA synthase (CBDAS), and CBCA synthase (CBCAS) then contribute to the diversity of cannabinoids (<a href=\"#figure1\">Figure 1</a>) [<a href=\"#r-13\">13</a>].</p>\r\n\r\n<p>Terpenes, vital compounds in plant biology, are synthesised through two distinct pathways. The cytosolic mevalonic acid (MVA) pathway produces sesqui- and triterpenes, while the plastid-localized MEP pathway synthesises mono-, di-, and tetraterpenes. These pathways start from acetyl-coenzyme A and pyruvate and d-glyceraldehyde-3-phosphate, respectively, leading to the production of isopentenyl diphosphate (IPP) and dimethylallyl diphosphate (DMAPP) as the end products. In the cytosol, farnesyl diphosphate synthase (FPS) combines two molecules of IPP (C5) and one molecule of DMAPP (C5) to form FPP, a precursor for sesquiterpenes (C15). Squalene synthase (SQS) in the endoplasmic reticulum uses two FPP to generate precursors for triterpenes and sterols. One molecule of IPP and one molecule of DMAPP are condensed to form an intermediate precursor of monoterpenes GPP (C10) by GPP synthase (GPS) in the plastid [<a href=\"#r-13\">13</a>] (<a href=\"#figure1\">Figure 1</a>).</p>\r\n\r\n<p>Phenolic compounds are synthesized in the cytoplasm through the phenylpropanoid pathway and transported to the vacuole or cell wall. The main classes of phenolic compounds are produced via the core phenylpropanoid pathway from phenylalanine to p-coumaroyl CoA, involving enzymes like phenylalanine-ammonia-lyase (PAL), cinnamate 4-hydroxylase (C4H, a cytochrome P450) and 4-coumarate-CoA ligase (4CL). Additional pathways lead to the formation of simple esters, lignins and lignans, flavonoids, coumarins, and stilbenes. The flavonoid pathway starts with condensing p-coumaroyl CoA and malonyl-CoA (<a href=\"#figure1\">Figure 1</a>), producing various flavonols like kaempferol and quercetin. In the next step, naringenin is converted to apigenin and luteolin, which are precursors of cannflavins (Figure 1) [<a href=\"#r-13\">13</a>].</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"655\" src=\"/media/article_images/2024/19/01/280-1711854419-Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1.</strong> Biosynthetic pathways of secondary metabolites production in <em>Cannabis sativa</em>. The cannabinoids, terpenoids, and flavonoids are produced in <em>Cannabis.</em> Glandular trichomes, located on the aerial parts and female flowers, serve as production and storage sites for cannabinoids and terpenoids. Cannabinoid synthesis involves olivetolic acid (OLA), from the polyketide pathway, and geranyl diphosphate (GPP), from the deoxyxylulose phosphate/methyl-erythritol phosphate (DOXP/MEP) pathway, as key precursors. Terpenoids, such as monoterpenoids, sesquiterpenoids, and triterpenoids are produced via the Plastidial mevalonate (MVA) or DOXP/MEP pathways (PMPP) or cytoplasmic mevalonate pathway (CMP). The main flavonoid classes, flavones and flavonols, are synthesized in <em>Cannabis sativa</em> through the phenylpropanoid pathway (PPP), converting phenylalanine to p-coumaroyl-CoA.</figcaption>\r\n</figure>\r\n</div>"
},
{
"section_number": 3,
"section_title": "LIMITATIONS OF CONVENTIONAL BREEDING",
"body": "<p><em>C. sativa</em> has nine pairs of autosomes and a pair of sex chromosomes (XY for males and XX for females). The determination of sex in <em>Cannabis</em> is influenced by several environmental factors. Sex chromosomes in <em>Cannabis</em> are not well understood, making sex determination a challenge in breeding new varieties. Dioecy in <em>Cannabis</em> is controlled by two specific genes at linked loci. Sex can only be determined at the beginning of flowering when male and female flowers are visible. So Dioecy caused heterogeneity is a crucial constraint in conventional breeding [<a href=\"#r-21\">21,22</a>].</p>\r\n\r\n<p>General <em>Cannabis</em> and other drug-type varieties have historically been bred through mass selection to improve quality traits like fiber, oil, and cannabinoid content. Early genetic improvement efforts faced challenges in avoiding high THC hemp genotypes and obtaining uniform medical genotypes. Recent advancements in breeding involve controlled mating of selected individuals from different landraces and cultivars. Synthetic varieties are created through open pollination using multiple female and male plants propagated via cuttings. High THC-type <em>Cannabis</em>, being dioecious, is often vegetatively propagated indoors to maintain genetic purity and uniformity. Indoor cultivators typically rely on cuttings from a mother plant, but this method requires space and can lead to decreased vigor, susceptibility to pests and diseases, and potential disease transmission [<a href=\"#r-21\">21, 22</a>].</p>\r\n\r\n<p><em>In vitro</em> techniques present a promising approach for large-scale production and germplasm maintenance. Tissue culture-based clean plant programs have successfully controlled plant pests, diseases, and viruses in vegetatively propagated crops and horticultural and ornamental crops. Developing an optimized <em>in vitro</em> method for propagating clean plants is crucial for producing genetically identical plants on a large scale, preserving genetic fidelity, and ensuring the long-term sustainability of economically significant <em>Cannabis </em>varieties<em> </em>[<a href=\"#r-14\">14</a>, <a href=\"#r-22\">22</a>].</p>"
},
{
"section_number": 4,
"section_title": "RECENT BIOTECHNOLOGICAL ADVANCES IN CANNABIS",
"body": "<p><strong><em>In vitro</em></strong><strong> plant<em> </em>growth and development</strong></p>\r\n\r\n<p>Recent advances of <em>Cannabis</em> biotechnology have led to improved plants with traits. Research on optimising <em>Cannabis</em> micropropagation has found that genotype, nutrient composition of the medium, and growth regulator supplementation significantly influence <em>Cannabis </em>propagation efficiency [<a href=\"#r-23\">23</a>, <a href=\"#r-24\">24</a>]. Establishing procedures for iterative micropropagation is essential for producing plant material for research purposes and multiplying germplasm efficiently [<a href=\"#r-25\">25</a>]. While MS-based media are commonly used for <em>C. sativa</em> micropropagation, some studies suggest that a DKW-based medium with increased levels of sulfur (~7×), calcium (~3×), and copper (10×) can enhance stage 2 explant growth over an extended period without decline [<a href=\"#r-26\">26</a>].</p>\r\n\r\n<p>Glucose has been tested as a carbon source for <em>in vitro</em> seed germination of industrial hemp. Still, it showed lower germination rates than sucrose, commonly used in hemp micropropagation studies. Glucose resulted in more vigorous plantlets across ten hemp genotypes, indicating an interaction between nutrient medium and carbon source for plant vigour. Maintaining a proper sugar-to-nitrate ratio is crucial for nitrogen uptake <em>in vitro</em>, with the highest ratios observed in DKW–glucose medium promoting vigorous growth [<a href=\"#r-27\">27</a>].</p>\r\n\r\n<p>Microbial contamination is a common issue in tissue culture due to nutrient-rich media commonly used Murashige and Skoog basal medium or its variants [28]. H<sub>2</sub>O<sub>2</sub> is effective against microbes [<a href=\"#r-29\">29</a>] and also plays a role in plant signaling for germination [<a href=\"#r-30\">30</a>]. A 1% H<sub>2</sub>O<sub>2 </sub>solution promoted rapid and successful germination of various <em>C. sativa</em> genotypes and contamination-free seedlings as explant sources in antibiotic-free MS media [<a href=\"#r-31\">31, 32</a>]. <em>In vitro</em>, <em>Cannabis</em> seed germination is slower than field germination due to the absence of microbes that aid seed coat digestion. Scarification or removal of the seed coat is crucial for breaking seed dormancy, promoting faster germination with sodium hypochlorite and hydrogen peroxide, and improving germination speed and frequency. Low salt concentrations enhanced seed germination rate and resulted in longer hypocotyls and radicles compared to high salt concentrations [<a href=\"#r-31\">31</a>, <a href=\"#r-33\">33</a>].</p>\r\n\r\n<p>The source of the explant is a crucial factor in rejuvenation practices to combat culture decline. Variability among explant types hinders the low efficiency of in vitro plant regeneration protocols for <em>C. sativa</em>. This variation poses a challenge for using tissue culture to improve this species. When two primordia emerge from the top of hypocotyls, they are always located on the organ's periphery and aligned in a specific way, leading to consistent plant regeneration from the same type of cells. This regeneration pattern closely resembles the regeneration area of 7 days seedling hypocotyl-derived meristems treated with ZEARIB 2.0 (mg/L) or ZEARIB 1.0 (mg/L) + NAA 0.02 (mg/L), resulting in a 66.67% response rate without any plant growth regulator application [<a href=\"#r-34\">34</a>]. The tissue's size, type, and developmental phase influence the success of response induction. Shoots originating from the basal and near-basal regions of the plant yielded taller shoots with more nodes than those from the middle and apical portions [<a href=\"#r-35\">35</a>]. Explant sources from basal portions yielded <em>Cannabis</em> plantlets characterised by shorter stature yet broader leaves, suggesting potential for <em>in vitro</em> rejuvenation with minimal culture degradation [<a href=\"#r-35\">35</a>].</p>\r\n\r\n<p>Hedging, a modified shoot tip culture technique, has been successfully applied in commercial horticultural crops [<a href=\"#r-79\">79</a>]. This method involves repeated harvests to eliminate the need for replanting, saving labour and resources. In <em>Cannabis</em> cultivation, an <em>in vitro</em> hedging technique was used to promote lateral shoot regrowth by removing shoot tips after three weeks [36]. The number of shoot tips harvested over multiple cycles, especially with higher light intensity. This method can enhance axillary divisions without the use of external plant growth regulators, leading to improved photoautotrophic metabolism [<a href=\"#r-35\">35, 36</a>]. Cuttings in the initial phases are placed in vessels with passive gas exchange under aseptic conditions, which is crucial for developing photoautotrophic micropropagation. In this technique, chlorophyllous explants are grown in CO<sub>2</sub>-rich environments. Using gas-permeable film vessels and porous substrate has proven effective for the photoautotrophic growth of plant species utilising a double-phase culture system, where shooting and rooting occur on semi-solid and liquid medium layers termed bioreactors. Such bioreactors have also been used to alter explant physiology. Rooting sponges have shown promising improvement in the aeration of the rhizosphere [<a href=\"#r-53\">53,54</a>].</p>\r\n\r\n<p>Plant cellular development encompasses the emergence of shoot primordia, apical meristems, leaf primordia, and procambium strands originating from the base of the floral explant where meristem centers develop at the junction between the filament and tepal, a phenomenon known as floral reversion [<a href=\"#r-37\">37</a>]. The mode of floral reversion is species-specific and holds significance in plant biotechnology based on transient gene expression have revealed the potential to express transgenes in floral tissues, rendering floral reversion a valuable tool in commercial crop production and for species that can overcome challenges in genetic modification [<a href=\"#r-38\">38</a>]. TDZ, a compound with auxin and cytokinin-like properties, enhanced callus induction in <em>C. sativa</em> floral tissues. Additionally, Using cytokinins, mT(metatopolin), and BAP promoted better shoot proliferation instead of callus induction from in pairs of florets rather than single florets [<a href=\"#r-38\">38</a>].Rooting and acclimatization are challenging stages in hemp and <em>Cannabis</em> tissue culture. Using sodium metasilicate, AgNO3, IBA, kinetin, or 2,4-D can aid this process. Sodium metasilicate enhances foliage appearance and rooting rate. The efficiency of plantlet recovery depends on <em>Cannabis</em> genotype, endophytic contamination, and rooting frequency [<a href=\"#r-39\">39</a>]. Incorporating potassium silicate as a silicon source into the growth medium demonstrated a notable reduction in hyperhydricity symptoms across three cultivars, concurrently enhancing root induction [<a href=\"#r-28\">28</a>].</p>\r\n\r\n<p>Low branching tendency and high apical dominance in fiber-type hemp result in a low multiplication rate and difficulties in initiating multi-shoot cultures [<a href=\"#r-24\">24</a>, <a href=\"#r-40\">40</a>]. Apical dominance is a plant phenomenon where the main shoot inhibits the growth of axillary buds [<a href=\"#r-41\">41</a>] and it is controlled by a complex network of hormones, including auxin flow from lateral buds [<a href=\"#r-42\">42-44</a>] and hormonal interactions between strigolactones and cytokinins play a role in bud outgrowth [<a href=\"#r-45\">45-47</a>]. The outgrowth of axillary buds depends on the ratio of these plant hormones [44, 46, 48]. The N-1-naphtylphtalamic acid, and 2,3,5-triiodobenzoic acid (TIBA) are known as auxin polar transport inhibitors that can break apical dominance and enhance shoot regeneration. These findings are valuable for the micropropagation of challenging industrial hemp varieties [<a href=\"#r-48\">49</a>].</p>\r\n\r\n<p>One approach to enhancing the rate of shoot multiplication <em>in vitro</em> is to interfere with apical dominance, which is often significant in <em>Cannabis</em> due to high levels of endogenous auxin. A promising compound for this purpose is α-(2-oxo-2-phenylethyl)-1H-indole-3-acetic acid (PEO-IAA), known for its solid anti-auxin activity. Auxin is a crucial plant hormone that regulates apical dominance through a short but universal signalling pathway that swiftly shifts between repressing and activating gene transcription by degrading transcriptional repressors dependent on auxin. At elevated auxin levels, auxin is believed to enhance binding between TIR1/AFB and AUX/IAA repressors, forming the core of the auxin signalling pathway long with auxin-responsive (ARF) transcription factors. When auxin levels drop, AUX/IAA repressors become more stable, increasing their interaction with ARF to suppress transcriptional activities. These auxin signalling cascades govern various cellular processes by activating or repressing distinct gene groups, including cell growth, expansion, and differentiation, which remain unclear. RT-qPCR analysis demonstrated that PEO-IAA in the culture medium affected the relative gene expression of targeted metabolites biosynthetic genes (OAC, CBCA, CBDA, THCA) and the levels of essential cannabinoids (CBCA, CBDA, CBC, ∆9-THCA, and ∆9-THC) [<a href=\"#r-50\">50, 51</a>].</p>\r\n\r\n<p><em>Cannabis</em> is typically a short-day plant, but auto-flowering genotypes are available. Short-day plants need a long dark period to induce flowering, which is more crucial than the light period [52]. Most <em>Cannabis</em> micropropagation is done under long-day photoperiods (16.0–18.0 hrs per day), keeping plants in a vegetative state. However, initial observations indicate that some genotypes flower <em>in vitro</em>, even under long days, showing the potential for <em>in vitro</em> flower development in <em>Cannabis</em>. Understanding genotype-specific photoperiods can optimize growth by providing the ideal conditions for photosynthesis. In a study, most <em>in vitro</em> flowering was seen under 12.0- and 13.2-hour treatments. <em>In vitro</em> flowering shows an efficient way to study floral and/or seed seed-based organ development and a new route of secondary metabolism in <em>Cannabis</em> [<a href=\"#r-53\">53</a>]. </p>\r\n\r\n<p><em>In vitro</em> culture can lead to genetic variation due to medium composition and plant growth regulators. The stability of regenerated plants is also a concern due to the frequent manipulation of different factors during micropropagation. To ensure the quality of plant material, genetic homogeneity was assessed using SSR markers in two <em>Cannabis</em> <em>sativa</em> varieties rich in CBD and CBG. This result suggested that <em>in vitro</em> multiplication protocols are suitable for clonal mass propagation with expected genetic stability [<a href=\"#r-54\">54</a>]. Overall discussions from the above, it can be concluded that complex interaction between several factors can make the <em>in vitro</em> growth and development of <em>Cannabis sativa</em> successful or unsuccessful. What they have yet to do is insufficient to claim their robustness. More focus should be given to multi-genotype trials for such outcomes.</p>\r\n\r\n<p><strong> </strong></p>\r\n\r\n<p><strong><em>In vitro</em></strong><strong> recalcitrance of <em>Cannabis </em>species</strong></p>\r\n\r\n<p>Various factors, such as plant hormones, nutrient composition, physical conditions, plant growth phase, explant nature, and season, are manipulated to induce organogenesis from the callus. However, despite these efforts, certain plant species are highly recalcitrant to tissue culture and may not induce organogenesis. The process of<em> </em>plant recalcitrance can inhibit plant organelle to response tissue manipulation [<a href=\"#r-55\">55</a>].</p>\r\n\r\n<p>A three-part regeneration system (callus-shooting-rooting) was previously successful in leaf explants from a high-THC genotype. It showed callogenesis on Murashige & Skoog (MS) medium with 0.5 μM of α-naphthaleneacetic acid (NAA) and 1.0 μM thidiazuron (TDZ) (Callus medium). Regeneration was achieved by transferring cultures to MS medium with 0.5 μM TDZ (shooting medium), resulting in a 96.6% response rate with an average of 12.3 shoots per culture. Rooting was successful on a medium with half-strength MS salts and 2.5 μM indole-3-butyric acid (IBA) (rooting medium). A multi-genotype study confirmed the effectiveness of the callus medium, achieving a 100% response rate across drug-type genotypes. These results highlight the importance of using multiple genotypes <em>in vitro</em> study with sufficient replication [<a href=\"#r-20\">20</a>]. <em>Cannabis </em>calli can be classified into two main groups, embryogenic and non-embryogenic, based on their morphology. The non-embryogenic callus can be friable or compact with no organ regeneration potential. Although not suitable for regeneration, it can be valuable for secondary metabolite production [<a href=\"#r-56\">56</a>].</p>\r\n\r\n<p>Embryogenic callus formation in <em>Cannabis sativa</em> is triggered by exogenous PGRs in tissue culture media. This process originates from cells with similar genetic backgrounds and morphologies. From a recent comparative transcriptomic study, upregulation of receptor-like proteins (RLPs) in embryogenic callus proved exogenous hormones play a vital role in <em>in vitro</em> regeneration. Most of the 12.4% of upregulated gene product properties were associated with cellular components. Cell wall modifications are essential for maintaining signalling factor transformation through a cellular connection. Various cell wall-modifying enzymes, cell wall-related signalling transduction genes, transcription factors interacting with chromatin modifiers (HDAC, CRF, etc.), Polycomb repressive complex (PRCs), and phytohormone-related genes differentially expressed in different types of calli, also highlighted the importance of PGRs in callogenesis. Studies have shown that imbalanced expression of auxin-cytokinin signalling pathway genes can lead to the conversion of non-embryogenic callus to embryogenic callus [<a href=\"#r-56\">56</a>]. The expression levels of genes involved in the cytokinin-dependent pathway resulted in embryogenic callus formation. In contrast, the repression of genes in the auxin-dependent pathway suppressed somatic embryogenesis. This suggests that the recalcitrant nature of <em>Cannabis</em> may be due to the repression of auxin-dependent pathway genes. Developing embryogenic tissues in <em>Cannabis</em> may differ from the conventional embryogenic pathway leading to plant regeneration. In this context, CRISPR-based genome editing techniques may provide exciting solutions for <em>Cannabis</em> development [<a href=\"#r-56\">56</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Temporary immersion system in <em>Cannabis sativa</em></strong></p>\r\n\r\n<p><em>In vitro,</em> micropropagation traditionally involves maintaining numerous culture vessels with semi-solid media and transferring plant material to fresh media every 4–6 weeks due to the medium's exhaustion of nutrients. This method is labour-intensive and costly due to the use of gelling agents. To reduce costs and improve efficiency, liquid medium and bioreactor cultures, such as Temporary Immersion system (TIS) for micropropagation, have become popular alternatives, allowing for easy plantlet scaling up, simplified handling, efficient nutrient uptake, and enhanced growth rates. However, challenges such as asphyxia, hyperhydricity, and physiological disorders may arise in liquid cultures [<a href=\"#r-57\">57, 58</a>]. Additionally, factors such as immersion frequency, liquid medium volume, number of explants, aeration, and forced ventilation are critical for optimising micropropagation using TIS [<a href=\"#r-58\">58, 59</a>].</p>\r\n\r\n<p>An efficient bioreactor approach with a ventilation system can improve photosynthesis and reduce contamination, leading to healthier shoots. <em>Cannabis </em>shoots in bioreactors grew with 0.5% sucrose but did not grow without sugar. CO<sub>2</sub>-enriched air can promote autotrophy. Bioreactor design influences micropropagation success. Future research could explore additional aeration in bioreactors, light intensity, light quality, Photoperiod, CO<sub>2</sub> supplementation, and sucrose’s impact on <em>Cannabis </em>rooting and acclimation in a variety of Bioreactor systems [<a href=\"#r-58\">58, 59</a>].</p>\r\n\r\n<p><strong> </strong></p>\r\n\r\n<p><strong><em>In vitro </em></strong><strong>polyploidization</strong></p>\r\n\r\n<p>Polyploidisation induced by antimitotic agents is common in plants and can increase genetic diversity in specific plant lineages. This process often results in polyploids with unique phenotypic traits, such as larger flowers or leaves. Studies have shown that tetraploids of medicinal plants, including <em>Cannabis</em>, exhibit higher concentrations of active metabolites [<a href=\"#r-60\">60, 61</a>]. <em>Cannabis</em> is a diploid plant with 20 chromosomes. Increasing the chromosome set through polyploidisation may enhance potency or customize cannabinoid ratios. Some studies suggest that polyploid <em>Cannabis</em> could have higher potency, but results vary, with some studies showing a decrease in THC levels.</p>\r\n\r\n<p>The polyploidisation on drug-type <em>Cannabis</em> strains is not well understood. In one study, a THC/CBD balanced drug-type strain of <em>C. sativa </em>was treated with the herbicide oryzalin to create polyploids. The specificity of oryzalin for plant tubulins is higher [<a href=\"#r-172\">172</a>] and is considered a more effective and less toxic alternative to colchicine. It also indicates that oryzalin is effective at over 30 times lower concentration than colchicine [<a href=\"#r-150\">150</a>,<a href=\"#r-153\">153</a>,<a href=\"#r-173\">173-176</a>].</p>\r\n\r\n<p>Cultured axillary bud explants were treated with various concentrations of oryzalin for 24 h, resulting in a higher number of tetraploids. Tetraploid plants exhibited larger leaves with leaves with 30% more stomata and 40% higher trichome density, critical sites for secondary metabolite production. There were observed considerable alterations in the terpene profile that influence the CBD levels in buds [<a href=\"#r-60\">60</a>, <a href=\"#r-62\">62</a>]. The cannabinoid ratio is influenced by specific enzymes on chromosome 6. Different cultivars have unique enzyme variants affecting cannabinoid production.</p>\r\n\r\n<p>Genome rearrangements like polyploidisation can create new enzyme combinations, leading to novel chemotypes with varied cannabinoid and terpene levels. Tetraploid sugar leaves show a 71.5% increase in terpene content due to higher trichome density. Buds also have about 30% more terpenes, indicating increased trichomes on flowers. However, the reason for the lack of a significant increase in cannabinoids with higher trichomes density remains unclear [<a href=\"#r-60\">60</a>, <a href=\"#r-61\">61</a>]. Additional analysis of tetraploid individuals in <em>Cannabis</em> can help to determine if polyploidisation results in larger floral size. Subsequent testing is needed to assess the stability of tetraploid clones over multiple generations and whether this stability is maintained when plants are propagated from seeds [<a href=\"#r-60\">60</a>, <a href=\"#r-63\">63</a>].</p>"
},
{
"section_number": 5,
"section_title": "IMPROVEMENT OF CANNABIS SATIVA",
"body": "<p><strong><em>In vitro</em></strong><strong> protoplast culture of <em>Cannabis sativa</em></strong></p>\r\n\r\n<p>Protoplasts are plant cells without cell walls, isolated from tissues like leaves, petals, and roots. They are totipotent and can form colonies, develop into callus, and regenerate plants. Protoplast fusion creates new cultivars with desired traits, especially in incompatible plants. Protoplasts can also be used for genetic modification, including CRISPR/Cas9-mediated genome editing. This technology allows for efficient validation of mutagenesis and DNA-free gene editing [<a href=\"#r-64\">64, 65</a>]. Hemp is a promising candidate for new plant breeding—technologies (NPBT) like CRISPR/Cas9-mediated gene editing. One potential target is the THCA synthase gene to create hemp plants with no THC production and potentially increased production of other significant minor cannabinoids. However, applying CRISPR/Cas9 to <em>Cannabis</em> breeding faces challenges due to the difficulty of working with <em>Cannabis</em> and the lack of methods for regenerating transgenic plants. Isolating protoplasts from <em>Cannabis</em> plants can be complex and influenced by species, growth conditions, tissue source, pretreatment, enzymes, and buffer composition (Figure 3) [<a href=\"#r-25\">25</a>].</p>\r\n\r\n<p>Auxin and its derivatives are frequently utilised in <em>Cannabis</em> tissue culture. Recent Studies have linked auxin signalling in <em>Cannabis</em> to the development of female flowers. Protoplasts co-transformed with the DR5: GFP reporter and auxin response factors, along with a plasmid containing a p35S: RFP expression cassette, showed successful transformation in 31% of protoplasts. These transformed protoplasts exhibited a robust response to a 16 h treatment with 5 μ M indole-3-acetic acid (IAA), resulting in nearly a four-fold increase in DR5:: GFP signal [<a href=\"#r-25\">25</a>]. </p>\r\n\r\n<p>Fluorescence microscopy analysis of the p35S: GFP construct showed that GFP signals of <em>CsCBCAS</em>, <em>CsCBDAS,</em> and <em>CsTHCAS</em> proteins were not evident as nuclear markers At ARR2-RFP. This suggests these proteins are localised in a cytoplasmic organelle outside the nucleus. However, the specific organelle where they are located remains unknown in <em>Cannabis</em> protoplasts. To determine the exact localisation of these enzymes, a co-expression study with cytoplasmic organelle markers for peroxisomes, mitochondria, vacuoles, and endoplasmic reticulum (ER) is required [<a href=\"#r-66\">66</a>].</p>\r\n\r\n<p>Optimising protoplast isolation from <em>Cannabis</em> involves a combination of digestion solutions. The best results were achieved with a protocol using 0.4M mannitol, a mix of enzymes (cellulase: macerozyme: pectolyase; 1.5%: 0.4%: 1.0), and vacuum-permeating treatment, yielding 9.7×10<sup>6</sup> viable protoplasts (g/ FLW). Enzymolysis times of 8 -16 h were optimal for high viability without over-digestion. Transient expression of GFP reached 23.2% efficiency with 30 μg plasmid, 50% PEG, 1 × 10<sup>6</sup> protoplasts, and a transfection duration of 20 min. [67, 68]. An investigation of what has been done till now is insufficient to establish a robust protocol. More studies are warranted to optimise all the factors for a thriving protoplast culture to manipulate metabolic pathway genes.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Improvement of <em>Cannabis </em>through genome editing</strong></p>\r\n\r\n<p>Genome editing is an efficient tool for introducing foreign genes into crops to enhance traits such as herbicide resistance, pro-vitamin A production, insect resistance, etc [<a href=\"#r-69\">69</a>]. In <em>Cannabis,</em> developing gene transformation and genome editing systems can modify horticultural traits, growth morphology, and stress resistance while also enabling gene function studies. <em>Agrobacterium </em>involving tumor-inducing gene delivery [<a href=\"#r-70\">70</a>] and <em>Agrobacterium</em>-mediated transformation is a common technique involving the transfer of genes via a plasmid containing phytohormone and opine synthase genes, which are transferred into plant cells through a type IV (pills) secretion system. Subsequently, they are incorporated into plant chromosomes, leading to gall formation. Recombinant strains can be created by engineering natural <em>A. tumefaciens </em>strains to delete genes within the T-DNA region from the Ti plasmid and insert genes of interest. Selectable marker genes, like those for antibiotic or herbicide resistance, are often used alongside genes for agronomic traits, like fungal disease resistance (<a href=\"#figure2\">Figure 2</a>) [<a href=\"#r-71\">71, 72</a>]. Explants were co-cultured with <em>A. tumefaciens</em> strain containing the neomycin phosphotransferase (nptII) genes. GUS histochemical analysis using the pBIN19 plasmid showed the vigor hypocotyl able to provide regeneration efficiency compared to cotyledons. Antibiotic phytotoxicity negatively affected explant regenerative ability during <em>Agrobacterium</em> co-culture, reducing plant regeneration rates for both hypocotyl and cotyledon of<em> C. sativa</em>. <em>Cannabis </em>seedlings showed less susceptibility to transformation by <em>A. tumefaciens</em> due to the influence of variety on susceptibility to <em>Agrobacterium</em> transformation treated with 1% H<sub>2</sub>O<sub>2 </sub>for seed sterilisation and germination [<a href=\"#r-73\">73</a>].</p>\r\n\r\n<p>Cutting leaf sections may induce secondary metabolite production, inhibiting <em>A. tumefaciens</em> growth and reducing TF (Transfer frequency). <em>Cannabis</em> extracts inhibited the development of 18 out of the 19 tested microbes. The inhibition of <em>Agrobacterium </em>growth by <em>C. sativa</em> metabolites must be confirmed and properly analysed to determine its impact on different <em>Cannabis</em> genotypes. [<a href=\"#r-74\">74</a>]. Non-expression of the pathogenesis-related protein 1 (NPR1) gene plays a crucial role in plants' salicylic acid-dependent pathway of systemic acquired resistance (SAR). Transformation with NPR1 or bar genes in <em>Agrobacterium</em> Strain EHA105 showed higher Transformation Frequency (TF) than strain GV3101. <em>Cannabis</em> genotypes likely influence transformation frequencies through their response to callus development [<a href=\"#r-74\">74</a>]. Excising <em>BBM</em> and <em>WUS</em> from transgenic plants is crucial because some adverse effects from these genes limit their application, although these two factors are significant for regeneration. Additional developmental genes like SHOOT MERISTEMLESS (STM) and ISOPENTENYL TRANSFERASE (IPT) induced shoot organogenesis. The GROWTH-REGULATING FACTOR (GRF) and GRF-INTERACTING FACTOR (GIF) transcriptional complex boosted genetic transformation in <em>Cannabis sativa,</em> proved through CRISPR/Cas9 technology [<a href=\"#r-75\">75</a>]. The susceptibility of genotypes to <em>Agrobacterium</em> has sparked interest in <em>Agrobacterium</em>-mediated gene transformation in <em>Cannabis.</em> However, challenges like low transformation efficiency, regeneration rates, chimeric regeneration, and transgene inactivation have been reported. To address these obstacles, it is essential to investigate factors like <em>Agrobacterium</em> strains, explants infection treatments, selection markers, chimerism eliminating, promoters, and translational enhancers [<a href=\"#r-15\">15</a>].</p>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"187\" src=\"/media/article_images/2024/19/01/280-1711854419-Figure2.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 2</strong>. The schematic representation of Agrobacterium-mediated gene transformation. In this process, the induction of de novo meristems (such as callus formation resembling in vitro plantlets, as well as callus-derived shoot and root regeneration) is essential. These steps are directly linked to plant tissue culture, which is critical for successful transfection.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"Table-1\">\r\n<p><a href=\"https://plant-trends.bsmiab.org/table/280-1711854419-table1/\">Table 1</a><strong>Table 1</strong>. Genetic manipulations in <em>Cannabis</em> <em>sativa</em> L.</p>\r\n</div>\r\n\r\n<p><strong> </strong></p>\r\n\r\n<p><strong>Hairy root culture of <em>Cannabis sativa</em></strong></p>\r\n\r\n<p>Limited information on the constituents and biological activities of hemp roots is available in the literature. Studies have shown that aqueous extract of the roots of <em>C. sativa</em> has anti-inflammatory and anti-asthmatic properties due to its unique active compounds. Hairy roots, which grow faster and produce valuable metabolites, can be induced with various agents to enhance the production of bioactive compounds [<a href=\"#r-81\">81</a>]. <em>Agrobacterium rhizogenes</em>, is a Gram-negative soil bacterium that creates hairy root syndromes as an output of hormonal changes. The infection begins with the bacterium moving towards wounded plant sites, attracted by simple phenolic compounds. <em>A. rhizogenes</em> then invades the plant tissue and transfers T-DNA from its Ri-plasmid using <em>virulence (vir)</em> genes. This T-DNA integrates into the plant genome, leading to hairy root development and opine synthesis for bacterial nutrition. Essential genes, such as root-inducing “rol” genes (<em>rolA, rolB, rolC, and rolD</em>), play a crucial role in modulating plant processes like phytohormone synthesis and metabolism. Among these, <em>rolB </em>is a significant regulator of secondary metabolism, activating specific transcription factors for metabolic pathways (<a href=\"#figure3\">Figure 3</a>) [<a href=\"#r-81\">81, 82</a>]. Triterpenoids have been identified in hemp roots, showing bioactive effects—the potential enhancement of triterpenoid accumulation in the roots of <em>C. sativa</em> through elicitation. Hairy roots were established successfully, exhibiting 2.02-fold higher triterpenoid content than natural roots. Elicited hairy roots with 75 μM salicylic acid showed 1.95-fold higher friedelin and epifriedelanol levels than untreated hairy roots. This suggested that the optimising elicitation of hairy root cultures could be a viable method for enhancing triterpenoid production. [<a href=\"#r-81\">81, 82</a>].Very few strains are tested for Hairy root culture-based Secondary metabolites production in <em>Cannabis sativa. </em>Future research should be focused on multi-strain trials available in Databases.</p>\r\n\r\n<div id=\"figure3\">\r\n<figure class=\"image\"><img alt=\"\" height=\"459\" src=\"/media/article_images/2024/19/01/280-1711854419-Figure3.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 3.</strong> <em>Agrobacterium rhizogenes</em> mediated genetic modification for improving plants and metabolite production. The infection process begins when the bacterium is attracted to a plant's wounded site through chemotaxis. It then penetrates the plant tissue, transferring TL-DNA (such as <em>rolA, rolB, rolC, and rolD</em> genes) into the plant genome. This transfer is facilitated by the virulence (<em>vir</em>) genes on the Ri-plasmid and the chv genes on the bacterial chromosomal DNA. Once integrated, the TL-DNA promotes the production of phytohormones like auxin and cytokinin, leading to the development of the characteristic hairy roots.</figcaption>\r\n</figure>\r\n</div>"
},
{
"section_number": 6,
"section_title": "ELICITATION IN CANNABIS SATIVA",
"body": "<p>Plants produce secondary metabolites to adapt to various stresses, which have industrial applications and potential for commercialisation. Plant cell/tissue culture is an alternative method for <em>in vitro</em> production of these metabolites, with elicitation being an essential technique. Different elicitors (e.g. abiotic or biotic) induce secondary metabolites, osmolytes, antioxidant enzymes as a response of plant defence mechanisms [<a href=\"#r-83\">83-87</a>]. However, the integrating transcriptomics, proteomics, and metabolomics with system biology can help discover new genes and pathways for metabolic engineering to increase secondary metabolite yield [<a href=\"#r-88\">88</a>].</p>\r\n\r\n<p>Elicitors interact with specific receptors on the plasma membrane, depolarizing and activating channels like K<sup>+</sup>/H<sup>+</sup> antiporter, causing cytoplasmic acidification and signalling for secondary metabolite production. Receptor perception can also trigger G-protein-linked receptors or the mitogen-activated protein kinase (MAPK) cascade, resulting in Ca<sup>2+</sup> fluxes and activation of intracellular processes. G-protein-linked receptors activate phospholipases C (PLC), phosphatidylinositol-4, and 5-bisphosphate (PIP2), leading to the production of secondary messengers like diacylglycerol (DAG) and inositol-1, 4,5-trisphosphate (IP3), which activates protein kinase C (PKC) and mobilizes Ca<sup>2+ </sup>ions. This process can generate reactive oxygen species (ROS) and regulate the expression of defence genes. Crosstalk between signalling pathways, such as jasmonic acid (JA) and ethylene (ET), can regulate gene expression against various es (<a href=\"#figure4\">Figure 4</a>) [<a href=\"#r-88\">88, 89</a>].</p>\r\n\r\n<p>In a study, CHT, SA, JA, and MeJA were tested, along with elicitation and precursor feeding during the exponential growth phase of cell suspensions. Secondary metabolites are primarily synthesised after cell growth enters the exponential phase, as plant cells need to adjust to the new environment during the lag phase. JA and MeJA treatments increase secondary metabolite accumulation but can inhibit growth by suppressing mitosis. Precursor feeding and elicitor treatments can enhance secondary metabolite synthesis by providing substrate biosynthesis pathways. A beneficial strategy involved MeJA and tyrosine (Tyr) precursor feeding, which increased the activity and expression of PAL and tyrosine aminotransferase (TAT). 1H-NMR analyses identified aromatic compounds, including 4-hydroxyphenylpyruvate (4-HPP), tyrosol, and 2,2-diphenyl-1-picrylhydrazyl (DPPH) [89]. Recent developments in the application of elicitors in coordinating antioxidant defence in various medicinal and other plants [<a href=\"#r-90\">90</a>]. However, elicitor responsive plant defence it is crucial for producing plant metabolites in Cannabis and other plant species.</p>\r\n\r\n<div id=\"figure4\">\r\n<figure class=\"image\"><img alt=\"\" height=\"397\" src=\"/media/article_images/2024/19/01/280-1711854419-Figure4.jpeg\" width=\"500\" />\r\n<figcaption><strong>Figure 4. </strong>Mechanism of elicitor-triggered signal transduction pathways leading to the synthesis of plant secondary metabolites. The elicitors, such as reactive oxygen species (ROS) and plant hormones, interact with specific receptors located on the plasma membrane, initiating multiple signaling pathways. Upon receptor activation, a rapid influx of calcium ions (Ca²⁺ burst) occurs, which is crucial for initiating intracellular signal transduction. The Ca²⁺ ions activate G-proteins, leading to downstream protein phosphorylation and mitogen-activated protein kinase (MAPK) activation. H₂O₂, a key reactive oxygen species, is generated and acts as a secondary messenger in elicitor-induced signaling, playing a role in the activation of calcium-dependent protein kinases (CDPKs), cyclic GMP (cGMP), cyclic AMP (cAMP), and phosphoinositide signaling pathways, which further enhance MAPK activation. Phospholipase C (PLC) is activated, resulting in the cleavage of phosphatidylinositol bisphosphate (PIP₂) to generate inositol trisphosphate (IP₃) and diacylglycerol (DAG). IP₃ mediates the release of Ca²⁺ from intracellular stores, reinforcing the calcium signaling cascade. Jasmonic acid, derived from linolenic acid, is also produced via this pathway and plays a role in the regulation of secondary metabolite production. The figure highlights the involvement of potassium (K⁺) and chloride (Cl⁻) ion fluxes, as well as the movement of hydrogen ions (H⁺) through ion channels, leading to cytoplasmic acidification. These processes are crucial for maintaining ion homeostasis and promoting the activation of downstream signaling pathways. The activation of MAPKs triggers a cascade of phosphorylation events, leading to the modulation of transcription factors (TFs) within the nucleus, which regulate the expression of genes involved in secondary metabolite biosynthesis. The coordinated activation of these signaling pathways culminates in the production of secondary metabolites, which play essential roles in plant defense and stress adaptation. Enzyme activation further enhances these biosynthetic processes.</figcaption>\r\n</figure>\r\n</div>"
},
{
"section_number": 7,
"section_title": "CONCLUSION AND FUTURE DIRECTIONS",
"body": "<p>This study provides an extensive overview of<em> </em>plant improvement and metabolite production in <em>Cannabis</em> through<em> </em>various biotechnological tools. Regarding its pharmaceutical and medicinal benefits, the plant has been improved for metabolite-containing traits using numerous biotechnological techniques. Significant advancements have been achieved through in vitro plant tissue culture, protoplast culture, genetic transformation, and modern gene editing approaches. These developments have greatly improved plant development, particularly enhanced growth with desired traits. The phytochemicals and bioactive compounds produced by <em>Cannabis </em>have significantly enriched its pharmaceutical and medicinal value globally. Despite these biotechnological and pharmaceutical advancements in <em>Cannabis</em>, some limitations still exist, which should be addressed and solved. However, several factors, especially the lack of robust genotype protocols, still show difficulties. Therefore, the growth and development of regulatory genes need to be studied with greater precision to produce transgenic <em>Cannabis</em> and enhance <em>in vitro</em> cell culture-based secondary metabolite production on an industrial scale. Furthermore, it is crucial to separate toxic or harmful metabolites, identify the factors involved in metabolite networks, and isolate novel genes encoding proteins or enzymes related to effective metabolite production in <em>Cannabis</em>. The modern CRISPR/Cas gene editing approach offers a potential solution to address these challenges in <em>C. sativa</em>.</p>"
},
{
"section_number": 8,
"section_title": "ACKNOWLEDGEMENTS",
"body": "<p>This work was supported by the Promotion (NO: P0023710) of Innovative Businesses for Regulation-Free Special Zones funded by the Ministry of SMEs and Startups (MSS, Korea).</p>"
},
{
"section_number": 9,
"section_title": "AUTHOR CONTRIBUTIONS",
"body": "<p>SMA and HWC conceived the research plan and wrote the initial draft of the manuscript. MIUH, SS, and MMR wrote and edited the manuscript. AK supported to figure drawing. All the authors approved the final version of the manuscript.</p>"
},
{
"section_number": 10,
"section_title": "CONFLICTS OF INTEREST",
"body": "<p>There is no conflict of interest among the authors.</p>"
}
],
"figures": [
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/19/01/280-1711854419-Figure1.jpg",
"caption": "Figure 1. Biosynthetic pathways of secondary metabolites production in Cannabis sativa. The cannabinoids, terpenoids, and flavonoids are produced in Cannabis. Glandular trichomes, located on the aerial parts and female flowers, serve as production and storage sites for cannabinoids and terpenoids. Cannabinoid synthesis involves olivetolic acid (OLA), from the polyketide pathway, and geranyl diphosphate (GPP), from the deoxyxylulose phosphate/methyl-erythritol phosphate (DOXP/MEP) pathway, as key precursors. Terpenoids, such as monoterpenoids, sesquiterpenoids, and triterpenoids are produced via the Plastidial mevalonate (MVA) or DOXP/MEP pathways (PMPP) or cytoplasmic mevalonate pathway (CMP). The main flavonoid classes, flavones and flavonols, are synthesized in Cannabis sativa through the phenylpropanoid pathway (PPP), converting phenylalanine to p-coumaroyl-CoA.",
"featured": false
},
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/19/01/280-1711854419-Figure2.jpg",
"caption": "Figure 2. The schematic representation of Agrobacterium-mediated gene transformation. In this process, the induction of de novo meristems (such as callus formation resembling in vitro plantlets, as well as callus-derived shoot and root regeneration) is essential. These steps are directly linked to plant tissue culture, which is critical for successful transfection.",
"featured": false
},
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/19/01/280-1711854419-Figure3.jpg",
"caption": "Figure 3. Agrobacterium rhizogenes mediated genetic modification for improving plants and metabolite production. The infection process begins when the bacterium is attracted to a plant's wounded site through chemotaxis. It then penetrates the plant tissue, transferring TL-DNA (such as rolA, rolB, rolC, and rolD genes) into the plant genome. This transfer is facilitated by the virulence (vir) genes on the Ri-plasmid and the chv genes on the bacterial chromosomal DNA. Once integrated, the TL-DNA promotes the production of phytohormones like auxin and cytokinin, leading to the development of the characteristic hairy roots.",
"featured": false
},
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/13/07/Figure_4.jpeg",
"caption": "Figure 4. Mechanism of elicitor-triggered signal transduction pathways leading to the synthesis of plant secondary metabolites. The elicitors, such as reactive oxygen species (ROS) and plant hormones, interact with specific receptors located on the plasma membrane, initiating multiple signaling pathways. Upon receptor activation, a rapid influx of calcium ions (Ca²⁺ burst) occurs, which is crucial for initiating intracellular signal transduction. The Ca²⁺ ions activate G-proteins, leading to downstream protein phosphorylation and mitogen-activated protein kinase (MAPK) activation. H₂O₂, a key reactive oxygen species, is generated and acts as a secondary messenger in elicitor-induced signaling, playing a role in the activation of calcium-dependent protein kinases (CDPKs), cyclic GMP (cGMP), cyclic AMP (cAMP), and phosphoinositide signaling pathways, which further enhance MAPK activation. Phospholipase C (PLC) is activated, resulting in the cleavage of phosphatidylinositol bisphosphate (PIP₂) to generate inositol trisphosphate (IP₃) and diacylglycerol (DAG). IP₃ mediates the release of Ca²⁺ from intracellular stores, reinforcing the calcium signaling cascade. Jasmonic acid, derived from linolenic acid, is also produced via this pathway and plays a role in the regulation of secondary metabolite production. The figure highlights the involvement of potassium (K⁺) and chloride (Cl⁻) ion fluxes, as well as the movement of hydrogen ions (H⁺) through ion channels, leading to cytoplasmic acidification. These processes are crucial for maintaining ion homeostasis and promoting the activation of downstream signaling pathways. The activation of MAPKs triggers a cascade of phosphorylation events, leading to the modulation of transcription factors (TFs) within the nucleus, which regulate the expression of genes involved in secondary metabolite biosynthesis. The coordinated activation of these signaling pathways culminates in the production of secondary metabolites, which play essential roles in plant defense and stress adaptation. Enzyme activation further enhances these biosynthetic processes.",
"featured": true
}
],
"authors": [
{
"id": 58,
"affiliation": [
{
"affiliation": "Department of Plant Medicals, Andong National University, Andong, 36729, Republic of Korea"
}
],
"first_name": "S.M.",
"family_name": "Ahsan",
"email": null,
"author_order": 1,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 13
},
{
"id": 59,
"affiliation": [
{
"affiliation": "Department of Applied Biosciences, Kyungpook National University, Daegu 41566, Republic of Korea"
}
],
"first_name": "Md. Injamum-Ul-",
"family_name": "Hoque",
"email": null,
"author_order": 2,
"ORCID": "http://orcid.org/0000-0003-3479-8105",
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 13
},
{
"id": 60,
"affiliation": [
{
"affiliation": "Department of Applied Biosciences, Kyungpook National University, Daegu 41566, Republic of Korea"
}
],
"first_name": "Ashim Kumar",
"family_name": "Das",
"email": null,
"author_order": 3,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 13
},
{
"id": 61,
"affiliation": [
{
"affiliation": "Department of Applied Biosciences, Kyungpook National University, Daegu 41566, Republic of Korea"
}
],
"first_name": "Shifa",
"family_name": "Shaffique",
"email": null,
"author_order": 4,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 13
},
{
"id": 62,
"affiliation": [
{
"affiliation": "Institute of Genomics for Crop Abiotic Stress Tolerance, Department of Plant and Soil Science, Texas Tech University, Lubbock, TX, 79409, USA"
}
],
"first_name": "Md. Mezanur",
"family_name": "Rahman",
"email": null,
"author_order": 5,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 13
},
{
"id": 63,
"affiliation": [
{
"affiliation": "Institute of Cannabis Biotechnology, Andong National University, Andong 36729, Republic of Korea"
},
{
"affiliation": "Department of Plant Medicals, Andong National University, Andong, 36729, Republic of Korea"
}
],
"first_name": "Hyong Woo",
"family_name": "Choi",
"email": "hwchoi@anu.ac.kr",
"author_order": 6,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": true,
"corresponding_author_info": "Hyong Woo Choi, PhD\r\nDepartment of Plant Medicals, Andong National University, Andong, 36729, Korea. Email: hwchoi@anu.ac.kr",
"article": 13
}
],
"views": 1425,
"downloads": 235,
"references": [
{
"id": 389,
"serial_number": 1,
"reference": "McPartland JM, Hegman W, Long T. Cannabis in asia: Its center of origin and early cultivation, based on a synthesis of subfossil pollen and archaeobotanical studies. Vegetation History and Archaeobotany. 2019;28:691-702.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 390,
"serial_number": 2,
"reference": "Sharma D, Bhushan S, et al. Cannabis as a potent therapeutic agent for pharmaceutical drugs: Recent advancement in drug discovery and human healthcare. In: Agrawal DC, Kumar R, Dhanasekaran M, editors. Cannabis/marijuana for healthcare. Singapore: Springer Nature Singapore; 2022. p. 77-99.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 391,
"serial_number": 3,
"reference": "Hesami M, Pepe M, et al. New insight into ornamental applications of cannabis: Perspectives and challenges. Plants. 2022;11:2383.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 392,
"serial_number": 4,
"reference": "Tanney CAS, Backer R, et al. Cannabis glandular trichomes: A cellular metabolite factory. Frontiers in Plant Science. 2021;12.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 393,
"serial_number": 5,
"reference": "Hesami M, Pepe M, et al. Recent advances in cannabis biotechnology. Industrial Crops and Products. 2020;158:113026.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 394,
"serial_number": 6,
"reference": "Shahbazi F, Grandi V, et al. Cannabinoids and cannabinoid receptors: The story so far. iScience. 2020;23:101301.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 395,
"serial_number": 7,
"reference": "Zou S, Kumar U. Cannabinoid receptors and the endocannabinoid system: Signaling and function in the central nervous system. International Journal of Molecular Sciences. 2018;19:833.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 396,
"serial_number": 8,
"reference": "Patel M, Finlay DB, Glass M. Biased agonism at the cannabinoid receptors – evidence from synthetic cannabinoid receptor agonists. Cellular Signalling. 2021;78:109865.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 397,
"serial_number": 9,
"reference": "Magham SV, Thaggikuppe krishnamurthy P, et al. Cannabinoid receptor 2 selective agonists and alzheimer's disease: An insight into the therapeutic potentials. Journal of Neuroscience Research. 2021;99:2888-905.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 398,
"serial_number": 10,
"reference": "Kovalchuk A, Wang B, et al. Fighting the storm: Could novel anti-tnfα and anti-il-6 c. Sativa cultivars tame cytokine storm in covid-19? Aging (Albany NY). 2021;13:1571-90.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 399,
"serial_number": 11,
"reference": "Nguyen LC, Yang D, et al. Cannabidiol inhibits sars-cov-2 replication through induction of the host er stress and innate immune responses. Science Advances. 2022;8:eabi6110.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 400,
"serial_number": 12,
"reference": "Onaivi ES, Sharma V. Cannabis for covid-19: Can cannabinoids quell the cytokine storm? Future Science OA. 2020;6.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 401,
"serial_number": 13,
"reference": "Andre CM, Hausman J-F, Guerriero G. Cannabis sativa: The plant of the thousand and one molecules. Frontiers in Plant Science. 2016;7.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 402,
"serial_number": 14,
"reference": "Adhikary D, Kulkarni M, et al. Medical cannabis and industrial hemp tissue culture: Present status and future potential. Frontiers in Plant Science. 2021;12.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 403,
"serial_number": 15,
"reference": "Hesami M, Baiton A, et al. Advances and perspectives in tissue culture and genetic engineering of cannabis. International Journal of Molecular Sciences. 2021;22:5671.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 404,
"serial_number": 16,
"reference": "Laverty KU, Stout JM, et al. A physical and genetic map of Cannabis sativa identifies extensive rearrangements at the thc/cbd acid synthase loci. Genome Research. 2019;29:146-56.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 405,
"serial_number": 17,
"reference": "Thomas F, Schmidt C, Kayser O. Bioengineering studies and pathway modeling of the heterologous biosynthesis of tetrahydrocannabinolic acid in yeast. Applied Microbiology and Biotechnology. 2020;104:9551-63.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 406,
"serial_number": 18,
"reference": "Bhajan S, Hasan M, et al. An efficient approach of in vitro plant regeneration and propagation o f mungbean [Vigna radiata (L.) wilczek]. Plant Trends.2(3):46-56.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 407,
"serial_number": 19,
"reference": "Niazian M. Application of genetics and biotechnology for improving medicinal plants. Planta. 2019;249:953-73.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 408,
"serial_number": 20,
"reference": "Monthony AS, Kyne ST, et al. Recalcitrance of Cannabis sativa to de novo regeneration; a multi-genotype replication study. PLOS ONE. 2021;16:e0235525.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 409,
"serial_number": 21,
"reference": "Wróbel T, Dreger M, et al. The application of plant in vitro cultures in cannabinoid production. Biotechnology Letters. 2018;40:445-54.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 410,
"serial_number": 22,
"reference": "Barcaccia G, Palumbo F, et al. Potentials and challenges of genomics for breeding cannabis cultivars. Frontiers in Plant Science. 2020;11.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 411,
"serial_number": 23,
"reference": "Page SRG, Monthony AS, Jones AMP. Dkw basal salts improve micropropagation and callogenesis compared to ms basal salts in multiple commercial cultivars of Cannabis sativa. bioRxiv. 2020:2020.02.07.939181.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 412,
"serial_number": 24,
"reference": "Wróbel T, Dreger M, et al. Modified nodal cuttings and shoot tips protocol for rapid regeneration of cannabis sativa l. Journal of Natural Fibers. 2022;19:536-45.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 413,
"serial_number": 25,
"reference": "Beard KM, Boling AWH, Bargmann BOR. Protoplast isolation, transient transformation, and flow-cytometric analysis of reporter-gene activation in cannabis sativa l. Industrial Crops and Products. 2021;164:113360.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 414,
"serial_number": 26,
"reference": "Monthony AS, Page SR, et al. The past, present and future of Cannabis sativa tissue culture. Plants. 2021;10:185.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 415,
"serial_number": 27,
"reference": "Halstead MA, Garfinkel AR, et al. Hemp growth in vitro and in vivo: A comparison of growing media and growing environments across 10 accessions. Hort Science. 2022;57:1041-7.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 416,
"serial_number": 28,
"reference": "Cassells AC. Pathogen and biological contamination management in plant tissue culture: Phytopathogens, vitro pathogens, and vitro pests. In: Loyola-Vargas VM, Ochoa-Alejo N, editors. Plant cell culture protocols. Totowa, NJ: Humana Press; 2012. p. 57-80.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 417,
"serial_number": 29,
"reference": "Weston RJ. The contribution of catalase and other natural products to the antibacterial activity of honey: A review. Food Chemistry. 2000;71:235-9.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 418,
"serial_number": 30,
"reference": "Wojtyla Ł, Lechowska K, et al. Different modes of hydrogen peroxide action during seed germination. Frontiers in Plant Science. 2016;7.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 419,
"serial_number": 31,
"reference": "Hesami M, Pepe M, et al. Modeling and optimizing in vitro seed germination of industrial hemp (cannabis sativa l.). Industrial Crops and Products. 2021;170:113753.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 420,
"serial_number": 32,
"reference": "Ahsan SM, Shin JH, Choi HW. Availability of hydrogen peroxide solutions as a germination liquid medium for contamination-free in vitro seedling development of Cannabis sativa. Horticultural Science and Technology. 2022;40:605-13.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 421,
"serial_number": 33,
"reference": "Pepe M, Hesami M, Jones AMP. Machine learning-mediated development and optimization of disinfection protocol and scarification method for improved in vitro germination of cannabis seeds. Plants. 2021;10:2397.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 422,
"serial_number": 34,
"reference": "Galán-Ávila A, García-Fortea E, et al. Development of a direct in vitro plant regeneration protocol from cannabis sativa l. Seedling explants: Developmental morphology of shoot regeneration and ploidy level of regenerated plants. Frontiers in Plant Science. 2020;11.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 423,
"serial_number": 35,
"reference": "Hesami M, Adamek K, et al. Effect of explant source on phenotypic changes of in vitro grown cannabis plantlets over multiple subcultures. Biology. 2023;12:443.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 424,
"serial_number": 36,
"reference": "Murphy R, Adelberg J. Physical factors increased quantity and quality of micropropagated shoots of Cannabis sativa L. In a repeated harvest system with ex vitro rooting. In Vitro Cellular & Developmental Biology - Plant. 2021;57:923-31.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 425,
"serial_number": 37,
"reference": "Zayed EMM, Zein El Din AFM, et al. Floral reversion of mature inflorescence of date palm in vitro. Annals of Agricultural Sciences. 2016;61:125-33.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 426,
"serial_number": 38,
"reference": "Monthony AS, Bagheri S, et al. Flower power: Floral reversion as a viable alternative to nodal micropropagation in cannabis sativa. In Vitro Cellular & Developmental Biology - Plant. 2021;57:1018-30.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 427,
"serial_number": 39,
"reference": "Holmes JE, Lung S, et al. Variables affecting shoot growth and plantlet recovery in tissue cultures of drug-type Cannabis sativa L. Frontiers in Plant Science. 2021;12.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 428,
"serial_number": 40,
"reference": "Smýkalová I, Vrbová M, et al. The effects of novel synthetic cytokinin derivatives and endogenous cytokinins on the in vitro growth responses of hemp (Cannabis sativa L.) explants. Plant Cell, Tissue and Organ Culture (PCTOC). 2019;139:381-94.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 429,
"serial_number": 41,
"reference": "Barbier FF, Dun EA, Beveridge CA. Apical dominance. Current Biology. 2017;27:R864-R5.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 430,
"serial_number": 42,
"reference": "Kebrom TH. A growing stem inhibits bud outgrowth – the overlooked theory of apical dominance. Frontiers in Plant Science. 2017;8.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 431,
"serial_number": 43,
"reference": "Bertheloot J, Barbier F, et al. Sugar availability suppresses the auxin-induced strigolactone pathway to promote bud outgrowth. New Phytologist. 2020;225:866-79.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 432,
"serial_number": 44,
"reference": "Prusinkiewicz P, Crawford S, et al. Control of bud activation by an auxin transport switch. Proceedings of the National Academy of Sciences. 2009;106:17431-6.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 433,
"serial_number": 45,
"reference": "Cheng ZJ, Wang L, et al. Pattern of auxin and cytokinin responses for shoot meristem induction results from the regulation of cytokinin biosynthesis by auxin response factor3 Plant Physiology. 2012;161:240-51.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 434,
"serial_number": 46,
"reference": "Luisi A, Lorenzi R, Sorce C. Strigolactone may interact with gibberellin to control apical dominance in pea (Pisum sativum). Plant Growth Regulation. 2011;65:415-9.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 435,
"serial_number": 47,
"reference": "Li M, Wei Q, et al. The effect of auxin and strigolactone on atp/adp isopentenyltransferase expression and the regulation of apical dominance in peach. Plant Cell Reports. 2018;37:1693-705.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 436,
"serial_number": 48,
"reference": "Müller D, Waldie T, et al. Cytokinin is required for escape but not release from auxin mediated apical dominance. The Plant Journal. 2015;82:874-86.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 437,
"serial_number": 49,
"reference": "Dreger M, Szalata M. The effect of tiba and npa on shoot regeneration of Cannabis sativa L. Epicotyl explants. Agronomy. 2022;12:104.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 438,
"serial_number": 50,
"reference": "Šenkyřík JB, Křivánková T, et al. Investigation of the effect of the auxin antagonist peo-iaa on cannabinoid gene expression and content in Cannabis sativa L. Plants under in vitro conditions. Plants. 2023;12:1664.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 439,
"serial_number": 51,
"reference": "Žukauskaitė A, Saiz-Fernández I, et al. New peo-iaa-inspired anti-auxins: Synthesis, biological activity, and possible application in hemp (Cannabis sativa L.) micropropagation. Journal of Plant Growth Regulation. 2023;42:7547-63.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 440,
"serial_number": 52,
"reference": "Lumsden, P.J., Vince-Prue, D. The perception of dusk signals in photoperiodic time-measurement. Physiol Plant. 1984;60:427-32.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 441,
"serial_number": 53,
"reference": "Moher M, Jones M, Zheng Y. Photoperiodic response of in vitro cannabis sativa plants. HortScience horts. 2021;56:108-13.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 442,
"serial_number": 54,
"reference": "Ioannidis K, Tomprou I, et al. Genetic evaluation of in vitro micropropagated and regenerated plants of cannabis sativa l. Using ssr molecular markers. Plants. 2022;11:2569.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 443,
"serial_number": 55,
"reference": "Patel P, Sarswat SK, Modi A. Chapter 12 - strategies to overcome explant recalcitrance under in vitro conditions. In: Chandra Rai A, Kumar A, et al., editors. Advances in plant tissue culture: Academic Press; 2022. p. 283-94.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 444,
"serial_number": 56,
"reference": "Hesami M, Pepe M, et al. Transcriptomic profiling of embryogenic and non-embryogenic callus provides new insight into the nature of recalcitrance in cannabis. International Journal of Molecular Sciences. 2023;24:14625.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 445,
"serial_number": 57,
"reference": "Murthy HN, Joseph KS, et al. Bioreactor systems for micropropagation of plants: Present scenario and future prospects. Frontiers in Plant Science. 2023;14.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 446,
"serial_number": 58,
"reference": "Mirzabe AH, Hajiahmad A, et al. Temporary immersion systems (tiss): A comprehensive review. Journal of Biotechnology. 2022;357:56-83.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 447,
"serial_number": 59,
"reference": "Rico S, Garrido J, et al. A temporary immersion system to improve cannabis sativa micropropagation. Frontiers in Plant Science. 2022;13.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 448,
"serial_number": 60,
"reference": "Parsons JL, Martin SL, et al. Polyploidization for the genetic improvement of Cannabis sativa. Frontiers in Plant Science. 2019;10.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 449,
"serial_number": 61,
"reference": "Gantait S, Mukherjee E. Induced autopolyploidy—a promising approach for enhanced biosynthesis of plant secondary metabolites: An insight. Journal of Genetic Engineering and Biotechnology. 2021;19:4.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 450,
"serial_number": 62,
"reference": "Madani H, Escrich A, et al. Effect of polyploidy induction on natural metabolite production in medicinal plants. Biomolecules. 2021;11:899.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 451,
"serial_number": 63,
"reference": "Bharati R, Fernández-Cusimamani E, et al. Oryzalin induces polyploids with superior morphology and increased levels of essential oil production in Mentha spicata L. Industrial Crops and Products. 2023;198:116683.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 452,
"serial_number": 64,
"reference": "Naing AH, Adedeji OS, Kim CK. Protoplast technology in ornamental plants: Current progress and potential applications on genetic improvement. Scientia Horticulturae. 2021;283:110043.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 453,
"serial_number": 65,
"reference": "Yue J-J, Yuan J-L, et al. Protoplasts: From isolation to crispr/cas genome editing application. Frontiers in Genome Editing. 2021;3.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 454,
"serial_number": 66,
"reference": "Kim AL, Yun YJ, et al. Establishment of efficient cannabis (Cannabis sativa L.) protoplast isolation and transient expression condition. Plant Biotechnology Reports. 2022;16:613-9.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 455,
"serial_number": 67,
"reference": "Matchett-Oates L, Mohamaden E, et al. Development of a robust transient expression screening system in protoplasts of cannabis. In Vitro Cellular & Developmental Biology - Plant. 2021;57:1040-50.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 456,
"serial_number": 68,
"reference": "Brandt KM, Gunn H, et al. A streamlined protocol for wheat (Triticum aestivum) protoplast isolation and transformation with crispr-cas ribonucleoprotein complexes. Frontiers in Plant Science. 2020;11.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 457,
"serial_number": 69,
"reference": "Tabatabaei I, Dal Bosco C, et al. A highly efficient sulfadiazine selection system for the generation of transgenic plants and algae. Plant Biotechnology Journal. 2019;17:638-49.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 458,
"serial_number": 70,
"reference": "Islam MS, Akter M, et al. Isolation of agrobacterium tumefaciens strains from crown gall sample of dicot plants in Bangladesh. Current Research in Bacteriology. 2010;3:27-36.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 459,
"serial_number": 71,
"reference": "Wally O, Jayaraj J, Punja ZK. Broad-spectrum disease resistance to necrotrophic and biotrophic pathogens in transgenic carrots (Daucus carota L.) expressing an arabidopsis npr1 gene. Planta. 2009;231:131-41.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 460,
"serial_number": 72,
"reference": "Silva KJP, Mahna N, et al. Npr1 as a transgenic crop protection strategy in horticultural species. Horticulture Research. 2018;5.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 461,
"serial_number": 73,
"reference": "Sorokin A, Yadav NS, et al. Transient expression of the β-glucuronidase gene in Cannabis sativa varieties. Plant Signaling & Behavior. 2020;15:1780037.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 462,
"serial_number": 74,
"reference": "Holmes JE, Punja ZK. Agrobacterium-mediated transformation of thc-containing Cannabis sativa L. Yields a high frequency of transgenic calli expressing bialaphos resistance and non-expressor of pr1 (npr1) genes. Botany. 2023;101:498-512.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 463,
"serial_number": 75,
"reference": "Zhang X, Xu G, et al. Establishment of an agrobacterium-mediated genetic transformation and crispr/cas9-mediated targeted mutagenesis in hemp (Cannabis sativa L.). Plant Biotechnology Journal. 2021;19:1979-87.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 464,
"serial_number": 76,
"reference": "Galán-Ávila A, Gramazio P, et al. A novel and rapid method for agrobacterium-mediated production of stably transformed cannabis sativa l. Plants. Industrial Crops and Products. 2021;170:113691.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 465,
"serial_number": 77,
"reference": "Zhang X, Xu G, et al. Establishment of an agrobacterium-mediated genetic transformation and crispr/cas9-mediated targeted mutagenesis in hemp (Cannabis sativa L.). Plant Biotechnol J. 2021;19:1979-87.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 466,
"serial_number": 78,
"reference": "Matchett-Oates L, Mohamaden E, et al. Development of a robust transient expression screening system in protoplasts of cannabis. In Vitro Cellular & Developmental Biology - Plant. 2021;57:1040 - 50.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 467,
"serial_number": 79,
"reference": "Beard KM, Boling AWH, Bargmann BOR. Protoplast isolation, transient transformation, and flow-cytometric analysis of reporter-gene activation in Cannabis sativa L. Industrial Crops and Products. 2021.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 468,
"serial_number": 80,
"reference": "Sorokin A, Yadav NS, et al. Transient expression of the β-glucuronidase gene in Cannabis sativa varieties. Plant Signaling & Behavior. 2020;15.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 469,
"serial_number": 81,
"reference": "Biswas D, Chakraborty A, et al. Hairy root culture: A potent method for improved secondary metabolite production of solanaceous plants. Frontiers in Plant Science. 2023;14. 1197555.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 470,
"serial_number": 82,
"reference": "Kobtrakul K, Rani D, et al. Elicitation enhances the production of friedelin and epifriedelanol in hairy root cultures of cannabis sativa l. Frontiers in Plant Science. 2023;14.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 471,
"serial_number": 83,
"reference": "Chauhan J, Srivastava JP, et al. Alterations of oxidative stress indicators, antioxidant enzymes, soluble sugars, and amino acids in mustard [Brassica juncea (L.) Czern and coss.] in response to varying sowing time, and field temperature. Frontiers in Plant Science. 2022;13. 875009.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 472,
"serial_number": 84,
"reference": "Raza A, Bhardwaj S, et al. Trehalose: A sugar molecule involved in temperature stress management in plants. The Crop Journal. 2024;12:1-16.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 473,
"serial_number": 85,
"reference": "Singh D, Sharma NL, et al. Exogenous hydrogen sulfide alleviates chromium toxicity by modulating chromium, nutrients and reactive oxygen species accumulation, and antioxidant defence system in mungbean (Vigna radiata L.) seedlings. Plant Physiology and Biochemistry. 2023;200:107767.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 474,
"serial_number": 86,
"reference": "Akash S, Etha S, et al. In vitro evaluation of antioxidant and antimicrobial properties of car dia mocleodii leaf extract. Plant Trends.2:38.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 475,
"serial_number": 87,
"reference": "Mahora M, Kigundu A, Waihenya R. Screening of phytochemicals, antioxidant activity, and in vivo safety profile of the hydroethanolic peel extract of musa sapientum. Plant Trends. 2(1):1-15.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 476,
"serial_number": 88,
"reference": "Narayani M, Srivastava S. Elicitation: A stimulation of stress in in vitro plant cell/tissue cultures for enhancement of secondary metabolite production. Phytochemistry Reviews. 2017;16:1227-52.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 477,
"serial_number": 89,
"reference": "Gabotti D, Locatelli F, et al. Cell suspensions of Cannabis sativa (var. Futura): Effect of elicitation on metabolite content and antioxidant activity. Molecules. 2019;24:4056.",
"pmc": null,
"DOI": null,
"article": 13
},
{
"id": 478,
"serial_number": 90,
"reference": "Hoque M, Jony M, et al. Coordination of elicitors and ascorbate-glutathione cycle: A vital nex us for mitigating post-harvest injury and losses of cultivated fruits. Plant Trends.2(2):24-37.",
"pmc": null,
"DOI": null,
"article": 13
}
]
},
{
"id": 12,
"slug": "280-1717504074",
"featured": false,
"slider": false,
"issue": "Vol2 Issue3",
"type": "original_article",
"manuscript_id": "280-1717504074",
"recieved": "2024-06-04",
"revised": null,
"accepted": "2024-08-07",
"published": "2024-08-31",
"pdf_file": "https://plant-trends.bsmiab.org/media/pdf_file/2024/18/280-1717504074.pdf",
"title": "Categorization of the flora and fauna: A study from the Uttara region, Dhaka, Bangladesh",
"abstract": "<p>Categorization of living things in a particular area is helpful to know their hypothetical relationships, similarities, dissimilarities, and present status of a particular species. The present study focused on a comprehensive taxonomic survey of the present floral arrangement in the semi-natural area of the Uttara region from August 2021 to October 2022. Basic methodological approaches and field surveys were employed to elucidate the floral diversity of the Uttara region, Dhaka, Bangladesh. The findings revealed a total of 225 plant species including, seasonal plants in the study area. These species were categorized into 119 families, with 39.11% of the species belonging to 43 families and the remaining 60.89% belonging to 76 different families. Among all families, Fabaceae is the largest family having 7% of the species. The rest of the plant families occupied 44% of species is the minor family having one species each. The recorded species were represented by trees (28.0 %), shrubs (16.0 %), herbs (43.55 %), and climbers (12.40 %). The plant species found in the study area were categorized into several functional groups. The entire population of plant communities was 225 including seasonal species, which were divided into 203 genera, 119 families, and 126, 46, 17, 15, 29, 10, and 14 plant species classified as ornamental, fruit, vegetable, spice, medicinal, timber, and plantation crop, respectively. To enhance the floral diversity and ecological balance of the area, it is necessary to implement sustainable management practices, public awareness, and participation in plant conservation activities, and should introduce sustainable native plant species. This study could be useful to plant taxonomists, plant ecologists, or naturalists for conducting region-specific surveys on the present status of species, or even flora and fauna.</p>",
"journal_reference": "Plant Trends. 2024; 2(3): 57-73",
"academic_editor": "AbdALmenem Hawamda, PhD; Palestine Technical University Kadoorie, Palestine",
"cite_info": "Naimuzzaman M, Chowdhury NQ, et al. Categorization of the flora and fauna: A study from the Uttara region, Dhaka, Bangladesh. Plant Trends. 2024; 2(3): 57-73.",
"keywords": [
"Flora",
"Plant resource",
"Uttara region",
"Plant diversity",
"Germplasm"
],
"DOI": "10.5455/pt.2024.06",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>Flora refers to the plant life or vegetation found in a particular region or ecosystem. It includes all plant species ranging from the smallest mosses and ferns to towering trees [<a href=\"#r-1\">1</a>]. Eventually, flora plays a vital role in the ecosystem, as it supports the food chain, contributes to the oxygen cycle, and helps to maintain the balance of nature. The study of flora, known as floristics, involves identifying and classifying plant species, studying their distribution and relationships, and understanding their ecological and evolutionary significance [<a href=\"#r-2\">2</a>]. Floral diversity plays a crucial role in maintaining the ecological balance of an ecosystem. Each plant species is part of a complex web of interactions between other species, including pollinators, herbivores, and predators [<a href=\"#r-3\">3</a>]. Unlike a wide variety of plants, the ecological system as a whole may go out of balance, which might have disastrous consequences for the environment and the organisms that rely on it [<a href=\"#r-4\">4</a>].</p>\r\n\r\n<p>Numerous attempts to study flowers have been made during the past few decades. Bangladesh has already conducted surveys and documented the plant species in on-premises areas of several universities [<a href=\"#r-5\">5-7</a>]. Although, the Uttara region displayed a diversification of plants. However, no floristic study was conducted on the Uttara region. With 165 million citizens, Bangladesh is among the most populous countries in the world [<a href=\"#r-8\">8</a>] and its population is now growing at a pace of 1.22%. Deforestation is a critical issue in Bangladesh. The country's forest cover has been drastically reduced over the past few decades due to logging, agricultural expansion, and illegal encroachments [<a href=\"#r-9\">9</a>]. The loss of forests not only diminishes biodiversity but also disrupts water cycles, increases soil erosion, and contributes to the loss of livelihoods for many communities dependent on forest resources [<a href=\"#r-10\">10</a>]. Carbon sequestration and nutrient dynamics constitute fundamental elements of ecosystem functionality [<a href=\"#r-11\">11</a>]. Plant communities act as carbon sinks, absorbing carbon dioxide from the atmosphere and mitigating the effects of climate change. In addition, diverse plant communities can help regulate local climates by providing shade, reducing temperatures, and maintaining humidity levels [<a href=\"#r-12\">12</a>]. In this scenario, plant resources are crucial for providing food, medicine, and other necessities. Nonetheless, it is regrettable that plant resources are disappearing globally, especially in Bangladesh (IUCN, 1990), and that this is a danger to farmland, the ecosystem, and the woodland [<a href=\"#r-13\">13</a>].</p>\r\n\r\n<p>In the center of Dhaka, Bangladesh, the Uttara region is a multicultural and active neighbourhood. The study area having incredible flower diversity, which gives the surroundings a special charm and beauty, is one of its most prominent qualities. The area is covered with a variety of trees, shrubs, and flowers that create a lush, green landscape and offer a welcoming setting for people. The goal of this research study is to examine the flora and fauna at the Uttara region by identifying the different plant species and exploring their ecological and monetary value. Hence, there is a high demand for enhanced knowledge in eco-friendly and cost-efficient methods to promote diverse plant population, maintain environment quality, and secure global food safety free from toxic substances [<a href=\"#r-14\">14</a>]. The study will also provide insight into how the locality protects and preserves its unique plant history, including how it uses sustainable plantations.</p>"
},
{
"section_number": 2,
"section_title": "MATERIALS AND METHODS",
"body": "<p><strong>Study Location</strong></p>\r\n\r\n<p>Uttara region is in the northern part of the capital of Bangladesh, Dhaka City at the bank of river Turag. Uttara region Uttara Thana (Dhaka metropolitan) area 36.91 sq km, located in between 23'51' and 23'52' north latitudes and in between 90'22' and 90'24' east longitudes. Of 9 m beyond the average sea level [<a href=\"#r-15\">15</a>]. The Uttara region Uttara Thana (Dhaka metropolitan) comprises an area of 36.91 sq km (<a href=\"#figure1\">Figure 1</a>). To guarantee the plant resources of plant communities at the Uttara region for this study, a field investigation was carried out.</p>\r\n\r\n<p><strong> </strong></p>\r\n\r\n<p><strong>Physiographic and soil condition of the study region</strong></p>\r\n\r\n<p>Uttara region lies under the Agro-ecological Zone - Modhupur Tract (AEZ 28), and it has a wet tropical climate having an average yearly temperature of 29.96°C (85.93°F), 9.9 inches of rainfall per year, and 65.8% mean yearly humidity [<a href=\"#r-16\">16</a>]. Reddish brown clay soil with a pH ranging from slightly acidic to extremely acidic makes up the soil types in these places. The soil analysis reveals that both the total content of one nutrient and the concentration of organic matter are at low to moderate levels. There are low levels of another nutrient and an essential trace element [<a href=\"#r-17\">17</a>, <a href=\"#r-18\">18</a>]. The study area is mentioned in <a href=\"#figure1\">Figure 1</a>.</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"386\" src=\"/media/article_images/2024/16/02/280-1717504074-Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1</strong>. The study area. A) Administrative units of Bangladesh with the location of Dhaka district, B) The administrative part of Dhaka City Corporation, C) Tejgaon – Uttara area and the study area Uttara region. The figure is developed using ArcMap 10.5.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong> Collection and compilation of data</strong></p>\r\n\r\n<p>A field survey was carried out (<a href=\"#Table-4\">Table 4</a>). The fallow land (plain), pond side bank, roadside, waterbody, roof-top, etc. were all recognized as potential flora habitats. Data were collected from August 2021 to October 2022. The study was of basic methodological approaches and surveys. Data were taken separately followed by local name, scientific name, area of collection, collection time, habit, habitat, family, and collector’s name attached with each data collection sheet. Most of these habitats were dispersed and intertwined throughout the study regions. We included every area that was accessible for the survey because the goal of the study was to find the full range of plant diversity in the study region. All kinds of witnessed plant species, including herbs, shrubs, trees, climbers, native and exotic species, cacti, orchids, and others, that were present in the habitats were recognized and named in the field.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Identification of the collected specimens and data analysis </strong></p>\r\n\r\n<p>With the use of published journals and textbooks of reference, Bengal Plants [<a href=\"#r-19\">19</a>], Encyclopedia of Flora and Fauna of Bangladesh [<a href=\"#r-20\">20</a>, <a href=\"#r-21\">21</a>], Trees of Bangladesh [<a href=\"#r-22\">22</a>], Medicinal Plants of Bangladesh [<a href=\"#r-23\">23</a>], and Red Data Book of Vascular Plants of Bangladesh [<a href=\"#r-24\">24</a>] the unidentified samples were recognized.</p>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p><strong>Number of plant species under different categories and habits</strong></p>\r\n\r\n<p>The comprehensive study of plant species in the Uttara region revealed a diverse floral community, categorized into <a href=\"#Table-1\">Table 1</a>, ornamental, fruit, vegetable, spice, medicinal, wood, plantation, and field crops, representing 39.11%, 16.44%, 16.0%, 4.44%, 12.44%, 2.22%, 3.11%, and 6.22%, respectively. Plants were further classified based on growth habits into trees (28.0%), shrubs (16.0%), herbs (43.55%), and climbers (12.44%). Ornamental plants predominated, followed by fruit and vegetable species, with timber plants showing the lowest density highlighting a trend towards urban gardening and ornamental horticulture [<a href=\"#r-25\">25</a>, <a href=\"#r-26\">26</a>].</p>\r\n\r\n<div id=\"Table-1\">\r\n<p><a href=\"https://plant-trends.bsmiab.org/table/280-1717504074-table1/\"><strong>Table 1</strong></a><strong>Table 1.</strong> List of total number of plant forms with their frequencies </p>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Distribution of family, genus, and species</strong></p>\r\n\r\n<p>The comprehensive documentation of plant species in the Uttara region highlights rich biodiversity, with a total of 225 plant species categorized into <a href=\"#Table-2\">Table 2</a>, ornamental, fruit, vegetable, spice, medicinal, timber, plantation, and field crops. The ornamental plants dominate, comprising 36.13% of the total families, followed by fruit plants at 20.16%, vegetables at 10.92%, spices at 5.04%, medicinal plants at 16.80%, timber at 3.36%, plantation crops at 1.68%, and field crops at 5.88%. In terms of genera, there are 89, 31, 25, 8, 25, 4, 7, and 14 for each respective category, and species count at 88, 37, 36, 10, 28, 5, 7, and 14. This diversity underscores the significance of ornamental plants in enhancing the campus landscape and supporting local biodiversity.</p>\r\n\r\n<div id=\"Table-2\">\r\n<p><a href=\"https://plant-trends.bsmiab.org/table/280-1717504074-table2/\">Table 2</a><strong>Table 2.</strong> List of family, genus and species with their frequencies</p>\r\n</div>\r\n\r\n<p><strong> </strong></p>\r\n\r\n<p><strong>Plant families with the percentage </strong></p>\r\n\r\n<p>The comprehensive documentation of plant species in the Uttara region stated in<a href=\"#figure2\"> Figure 2</a> reveals Fabaceae as the largest plant family, comprising 7% of all species. Following Fabaceae, Amaryllidaceae accounts for 4%, making it the second-largest plant family, with Cucurbitaceae, Moraceae, Rutaceae, and Solanaceae also contributing significantly to the floral diversity. Families such as Brassicaceae, Compositae, Euphorbiaceae, and Malvaceae each represent 3% of the total plant families, while Apocynaceae, Caesalpiniaceae, Combretaceae, Lamiaceae, Liliaceae, Palmae, Rubiaceae, and Zingiberaceae each account for 2%. The remaining 44% of plant families are represented by single species, highlighting a considerable presence of rare or less common families. This extensive diversity aligns with previous studies [<a href=\"#r-27\">27</a>, <a href=\"#r-28\">28</a>], emphasizing Fabaceae's ecological and agricultural significance, particularly in nitrogen fixation and soil fertility [<a href=\"#r-28\">28</a>, <a href=\"#r-29\">29</a>].</p>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"308\" src=\"/media/article_images/2024/16/02/280-1717504074-Figure2.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 2</strong>. The distribution (%) of studied plant families from the Uttara, region, Bangladesh. Different colours on Pie chart indicate family-based plant distribution (%), and right panel shows the identified plant families in alphabetic order.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Fruit plant species under each family</strong></p>\r\n\r\n<p>The study recorded in <a href=\"#Table-3\">Table 3</a> a total of 37 fruit tree species from 23 families in the Uttara region. The Rutaceae family had the highest number of species, with 6 species accounting for 16.21% of all fruit-producing plants. The Moraceae family followed with 4 species, representing 10.81% of the fruit tree species, while the Myrtaceae and Anacardiaceae families each contributed 3 species, making up 8.10% of the total fruit tree species. In contrast, only one species was found in several families, including Annonaceae, Caricaceae, Malvaceae, and Sapotaceae. These findings highlight the dominance of the Rutaceae family, consistent with its known diversity and economic importance, particularly in the cultivation of citrus fruits.</p>\r\n\r\n<div id=\"Table-3\">\r\n<p><a href=\"https://plant-trends.bsmiab.org/table/280-1717504074-table3/\">Table 3</a><strong>Table 3.</strong> List of fruit plant species with their families and frequencies</p>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<p><strong>List of flora </strong><strong>of Uttara region with their respective common names, scientific names, family, and plant type</strong></p>\r\n\r\n<p>Plant genetic resources represent a wide range of plant species essential for upholding global food security and agricultural resilience [<a href=\"#r-30\">30</a>]. The extensive variety of plant genetic resources highlights their importance in agricultural research, breeding initiatives, and conservation endeavors focused on preserving biodiversity and promoting agricultural sustainability [<a href=\"#r-31\">31</a>]. The enumeration of the botanical species presented in the Uttara region, inclusive of their corresponding colloquial nomenclature, taxonomic designations, families of various fauna specimens stated in <a href=\"#figure3\">Figure 3</a>, and plant type-based categorization was presented in <a href=\"#Table-4\">Table 4</a>.</p>\r\n\r\n<div id=\"figure3\">\r\n<figure class=\"image\"><img alt=\"\" height=\"379\" src=\"/media/article_images/2024/16/02/280-1717504074-Figure3.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 3</strong>. Photographs show various flora specimens and their corresponding scientific names. The plant species are identified from the Uttara region, Dhaka Bangladesh.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<div id=\"Table-4\">\r\n<p><a href=\"https://plant-trends.bsmiab.org/table/280-1717504074-table4/\">Table 4</a><strong>Table 4.</strong> List of the identified plant specimens in the Uttara region</p>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Ornamental plant species under each family</strong></p>\r\n\r\n<p>This study recorded a total of 88 ornamental plant species from 43 families in the Uttara region. The Compositae family had the highest number of species, with 7 species making up 7.95% of all ornamental plants. Following Compositae, the Apocynaceae family comprised 6 species, contributing 6.81% to the ornamental plant diversity.</p>\r\n\r\n<div id=\"Table-5\">\r\n<p><a href=\"https://plant-trends.bsmiab.org/table/280-1717504074-table5/\">Table 5</a><strong>Table 5.</strong> List of ornamental plant species with their families and frequencies</p>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Vegetable plant species under each family</strong></p>\r\n\r\n<p>The examination of vegetable plant species at Uttara region revealed a total of 37 species classified into 13 families, highlighting the significant diversity present stated in <a href=\"#Table-6\">Table 6</a>. The Cucurbitaceae family emerged as the most abundant, containing 9 species, which constitutes 24.32% of the overall vegetable plant species. Following closely, the Amaranthaceae family ranked second with 7 species, underscoring its importance in providing nutrient-rich leafy vegetables essential for food security.</p>\r\n\r\n<div id=\"Table-5\">\r\n<p><a href=\"https://plant-trends.bsmiab.org/table/280-1717504074-table6/\">Table 6</a><strong>Table 6.</strong> List of vegetable plant species with their families and frequencies</p>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Spices of plant and field crop species under each family</strong></p>\r\n\r\n<p>Six families of spice plants included a maximum of 11 species in <a href=\"#Table-7\">Table 7</a>. The greatest number of species were found in the Zingiberaceae family, accounting for 3 and 27.27% of all spice plants, respectively (<a href=\"#Table-7\">Table 7</a>). Amaryllidaceae, Lauraceae, and Solanaceae together make up 18.18% of all species of spice plants. A minimum of one plant species from the Rutaceae and Apiaceae. This study identified a total of 14 field crop species across seven families, with the Fabaceae family leading the count with six species, representing 42.85% of all field crops. Following closely, the Poaceae family contributed three species, accounting for 21.42%. The remaining families, including Cruciferae, Lamiaceae, Pedaliaceae, Solanaceae, and Tiliaceae, had minimal representation, collectively making up only 7.14% of the total. In parallel, the study also identified 11 species of spice plants from six families, where the Zingiberaceae family, with three species, constituted 27.27% of all spice plants.</p>\r\n\r\n<div id=\"Table-7\">\r\n<p><a href=\"https://plant-trends.bsmiab.org/table/280-1717504074-table7/\">Table 7</a><strong>Table 7.</strong> List of plants and field crop species with their families and frequencies</p>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Medicinal plant species under each family</strong></p>\r\n\r\n<p>This study documented in <a href=\"#Table-8\">Table 8</a> a total of 29 medicinal plant species across 20 families, with the Lamiaceae family showcasing the highest diversity, contributing four species, which represents 10.50% of all medicinal plants. The Combretaceae family followed, with three species accounting for 10.34% of the total. Families such as Acanthaceae, Asteraceae, Fabaceae, and Malvaceae each contributed two species, collectively making up 6.89% of the total. Individual species were also identified from families including Annonaceae, Boraginaceae, Cucurbitaceae, Dioscoreaceae, Euphorbiaceae, Labiatae, and Zingiberaceae, highlighting a diverse array of medicinal resources <a href=\"#Table-8\">Table 8</a>.</p>\r\n\r\n<div id=\"Table-8\">\r\n<p><a href=\"https://plant-trends.bsmiab.org/table/280-1717504074-table8/\">Table 8</a><strong>Table 8.</strong> List of medicinal plant species with their families and frequencies</p>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Timber plant and plantation crop species of Uttara region, Dhaka under each family</strong></p>\r\n\r\n<p>A maximum of 5 spice plants was seen in 4 families of trees. The greatest number of species discovered belonged to the Mimosaceae family, which made up 2 and 40% of all woody plants, respectively. The Boraginaceae family, followed by the Fabaceae and Maliaceae, account for 20% of all species of timber plants, which is the least amount <a href=\"#Table-9\">Table 9</a>. The study identified a total of seven spice plant species across six families, with the Palmae family exhibiting the highest diversity, comprising four species, which account for 57.14% of all plantation crops. The Arecaceae family followed closely with three species, representing 42.86% of the total. Additionally, five spice plant species were documented across four families of trees, where the Mimosaceae family emerged as the most prevalent, encompassing two species and making up 40% of the woody plants. The Boraginaceae, Fabaceae, and Maliaceae families contributed one species each, together constituting 60% of the overall timber plant species <a href=\"#Table-9\">Table 9</a>.</p>\r\n\r\n<div id=\"Table-9\">\r\n<p><a href=\"https://plant-trends.bsmiab.org/table/280-1717504074-table9/\">Table 9</a><strong>Table 9.</strong> List of timber plant species with their families and frequencies</p>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<p><strong>List of Fauna identified from the study area</strong></p>\r\n\r\n<p>The present survey findings displayed a variety of insect pests that were observed during the investigation <a href=\"#Table-10\">Table 10</a>. These pests appeared from several orders and families, highlighting the wide variety of insects that can endanger plants and crops. The list starts with widespread insects including White Fly (<em>Trialeurodes vaporariorum</em>) and Mealybug (<em>Ferrisia virgata</em>), that are infamous for infesting different plants and weakening them by consuming sap. The Leaf miner (<em>Tuta absoluta</em>), with larvae we found that tunnel through foliage and cause damage and decreased plant vitality is another serious pest. The commonly seen Common Fruit Fly (Drosophila melanogaster) is a noticeable presence because it contaminates ripening fruit, causing deterioration and monetary losses. Pests like Field Cricket (<em>Gryllus</em> sp.), Yellow stem borer (<em>Scirpophaga incertulas</em>), and Brown plant hopper (<em>Nilaparvata lugens</em>), which eat plant tissues or spread disease, can seriously harm crops like rice, also prevail in the present study. The survey revealed the pests unique to certain crops, such as the Rice root weevil (<em>Sitophilus oryzae</em>) and Rice gundhi bug (<em>Leptocorisa</em> spp.), which can severely harm rice grains. The Giant water bug (<em>Lethocerus americanus</em>), albeit not a significant agricultural pest, was also described as being present in rice fields.</p>\r\n\r\n<div id=\"Table-10\">\r\n<p><a href=\"https://plant-trends.bsmiab.org/table/280-1717504074-table10/\">Table 10</a><strong>Table 10.</strong> List of fauna identified in the Uttara region, Dhaka, Bangladesh.</p>\r\n</div>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p>The floristic survey of Uttara region reveals a rich and diverse array of plant species, underscoring the ecological and socio-economic significance of urban green spaces. This study identified 225 plant species across 119 families, demonstrating a considerable botanical variety that can significantly contribute to urban biodiversity, ecological resilience, and the local economy. The presence of ornamental, fruit, and medicinal plants highlights their role in enhancing urban biodiversity and providing essential ecosystem services. Ornamental plants, such as those from the Compositae and Apocynaceae families, add aesthetic value and support urban wildlife by providing habitats and food sources for various pollinators. Integrating diverse plant species in urban planning can enhance ecological balance, mitigate urban heat island effects, and improve air quality.</p>\r\n\r\n<p>The lower density of timber plants, reflective of urbanization and changing land use priorities, emphasizes the need for sustainable urban development that incorporates diverse plant species to maintain ecological resilience and promote biodiversity in densely populated areas like Dhaka [<a href=\"#r-32\">32</a>]. The presence of key families like Moraceae, Myrtaceae, and Anacardiaceae underscores their ecological and agricultural value, contributing to ecosystem services and biodiversity [<a href=\"#r-33\">33</a>, <a href=\"#r-34\">34</a>]. The substantial presence of fruit and medicinal plants indicates significant opportunities for educational and research endeavors in horticulture and pharmacognosy. Families such as Amaryllidaceae and Brassicaceae are notable for their diverse applications in food production and traditional medicine. For instance, plants from the Moringaceae family, including <em>Moringa oleifera</em>, are celebrated for their therapeutic properties, such as anti-diabetic, anti-inflammatory, and antioxidant effects [<a href=\"#r-35\">35</a>]. Similarly, the Combretaceae family, with plants like Terminalia, is known for its potent antioxidant and antibacterial compounds [<a href=\"#r-36\">36</a>]. These findings highlight the potential for utilizing these plant resources in developing new pharmaceuticals and enhancing nutritional health.</p>\r\n\r\n<p>The variety of vegetable and spice plants identified in the study offers opportunities for culinary and nutritional studies, contributing to dietary health and agricultural economics. The prominence of families such as Solanaceae and Amaryllidaceae, which include staples like chili peppers and garlic, underscores their importance in local cuisine and food security [<a href=\"#r-37\">37</a>]. Additionally, the presence of underutilized crops from families like Apiaceae and Dioscoreaceae suggests potential for expanding crop diversity and improving agricultural resilience [<a href=\"#r-38\">38</a>]. The representation of timber and plantation crops points to avenues for agroforestry research. The substantial presence of the Palmae family, with economically important plants like coconut and oil palm, underscores their vital role in agriculture and economic development [39]. These findings align with previous studies emphasizing the role of plant diversity in promoting sustainable agricultural practices and supporting local economies.</p>\r\n\r\n<p>The discovery of numerous rare plant families enhances overall biodiversity, essential for ecological resilience and research opportunities. The minimal representation of families like Annonaceae and Caricaceae suggests areas for potential expansion in cultivation and research, as these families may include species with unique traits beneficial for diversifying fruit production and improving crop resilience [<a href=\"#r-40\">40</a>]. This detailed inventory of plant species serves as a foundation for future studies aimed at optimizing urban horticulture and promoting sustainable agricultural practices. Continued research and monitoring are essential to understand the dynamics of plant populations and their interactions with urban environments. Implementing conservation strategies and sustainable management practices can help preserve plant diversity and ensure the long-term ecological health of urban areas.</p>\r\n\r\n<p>The report also highlights the prevalence of various insect pests that pose significant threats to crop production. Effective pest management strategies are crucial to protect plant productivity and minimize financial losses. The diversity of pests identified, including the whitefly, mealybug, and rice green leafhopper, underscores the need for targeted control measures and integrated pest management practices. By focusing on pests that affect multiple crops, the study emphasizes the broader challenge of managing pest populations across various agricultural sectors. Implementing effective pest control measures can enhance crop yields, reduce economic losses, and contribute to sustainable agricultural practices.</p>"
},
{
"section_number": 5,
"section_title": "CONCLUSION",
"body": "<p>The current study found that there are 225 different plant species, 119 families, and 203 different genera present in Uttara region. A total of 88 species of ornamental was categorized into 89 genera and 43 families (one was rare and endangered). Spice plants were displayed in ten species, with eight genera and six families each. Fruits were divided into 37 families, 37 genera, and 37 species. There were 36 species of vegetables divided into 25 genera and 13 families. A total of 5 species of timber plants were grouped into 4 genera and 4 families. Twenty families and twenty genera made up the 28 medicinal species. The plantation crops included 7 species, 7 genera, and 2 families. 14 species and 14 families made up the field crop. Many plant categories have a variety of behaviors, as well as a variety of species, genera, and families, and some insect pests are also observed. The variety and quantity of plant species in the study area showed a strong floristic diversity. The campus area has the potential to become a premier hub for the preservation of biological variety with the right management tactics, adoption of crucial laws, and conservation activities. This study offers important insights into the floral variety of the Uttara region and sets the groundwork for future ecological studies and conservation efforts.</p>"
},
{
"section_number": 6,
"section_title": "AUTHORS CONTRIBUTION",
"body": "<p>SKR and MN conceived the research plan and improved the methodologies. NQC, ZSA, SM, JS, JN, IAI, and FA contributed to the survey. MN and FR conducted the data analysis and table preparation and wrote the manuscript. SKR, FB, PKM, and MOS did the manuscript review and edited the manuscript. All the authors approved the final version of the manuscript.</p>\r\n\r\n<p> </p>"
},
{
"section_number": 7,
"section_title": "ACKNOWLEDGEMENT",
"body": "<p>The authors are grateful to the Bangladesh Forest Research Institute and the International University of Business Agriculture, and Technology (IUBAT).</p>"
},
{
"section_number": 8,
"section_title": "CONFLICTS OF INTEREST",
"body": "<p>There is no conflict of interest among the authors.</p>"
}
],
"figures": [
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/16/02/280-1717504074-Figure1.jpg",
"caption": "Figure 1. The study area. A) Administrative units of Bangladesh with the location of Dhaka district, B) The administrative part of Dhaka City Corporation, C) Tejgaon – Uttara area and the study area Uttara region. The figure is developed using ArcMap 10.5.",
"featured": false
},
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/16/02/280-1717504074-Figure2.jpg",
"caption": "Figure 2. The distribution (%) of studied plant families from the Uttara, region, Bangladesh. Different colours on Pie chart indicate family-based plant distribution (%), and right panel shows the identified plant families in alphabetic order.",
"featured": false
},
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/16/02/280-1717504074-Figure3.jpg",
"caption": "Figure 3. Photographs show various flora specimens and their corresponding scientific names. The plant species are identified from the Uttara region, Dhaka Bangladesh.",
"featured": true
}
],
"authors": [
{
"id": 45,
"affiliation": [
{
"affiliation": "College of Agricultural Sciences, International University of Business Agriculture and Technology, Dhaka-1230, Bangladesh"
}
],
"first_name": "Mollah",
"family_name": "Naimuzzaman",
"email": null,
"author_order": 1,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 12
},
{
"id": 46,
"affiliation": [
{
"affiliation": "College of Agricultural Sciences, International University of Business Agriculture and Technology, Dhaka-1230, Bangladesh"
}
],
"first_name": "Nafisa Quader",
"family_name": "Chowdhury",
"email": null,
"author_order": 2,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 12
},
{
"id": 47,
"affiliation": [
{
"affiliation": "College of Agricultural Sciences, International University of Business Agriculture and Technology, Dhaka-1230, Bangladesh"
}
],
"first_name": "Zizan Sharrad",
"family_name": "Anusha",
"email": null,
"author_order": 3,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 12
},
{
"id": 48,
"affiliation": [
{
"affiliation": "College of Agricultural Sciences, International University of Business Agriculture and Technology, Dhaka-1230, Bangladesh"
},
{
"affiliation": "Farzeen Biotechnology and Genetic Engineering Research Laboratory, International University of Business Agriculture and Technology, Dhaka-1230, Bangladesh"
}
],
"first_name": "Faizur",
"family_name": "Rahman",
"email": null,
"author_order": 4,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 12
},
{
"id": 49,
"affiliation": [
{
"affiliation": "College of Agricultural Sciences, International University of Business Agriculture and Technology, Dhaka-1230, Bangladesh"
}
],
"first_name": "Sajib",
"family_name": "Mia",
"email": null,
"author_order": 5,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 12
},
{
"id": 50,
"affiliation": [
{
"affiliation": "College of Agricultural Sciences, International University of Business Agriculture and Technology, Dhaka-1230, Bangladesh"
}
],
"first_name": "Jakia",
"family_name": "Sultana",
"email": null,
"author_order": 6,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 12
},
{
"id": 51,
"affiliation": [
{
"affiliation": "College of Agricultural Sciences, International University of Business Agriculture and Technology, Dhaka-1230, Bangladesh"
}
],
"first_name": "Jannatun",
"family_name": "Naim",
"email": null,
"author_order": 7,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 12
},
{
"id": 52,
"affiliation": [
{
"affiliation": "College of Agricultural Sciences, International University of Business Agriculture and Technology, Dhaka-1230, Bangladesh"
}
],
"first_name": "Istiaque Ahmed",
"family_name": "Isti",
"email": null,
"author_order": 8,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 12
},
{
"id": 53,
"affiliation": [
{
"affiliation": "College of Agricultural Sciences, International University of Business Agriculture and Technology, Dhaka-1230, Bangladesh"
}
],
"first_name": "Faiyaz",
"family_name": "Ahmed",
"email": null,
"author_order": 9,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 12
},
{
"id": 54,
"affiliation": [
{
"affiliation": "College of Agricultural Sciences, International University of Business Agriculture and Technology, Dhaka-1230, Bangladesh"
},
{
"affiliation": "Farzeen Biotechnology and Genetic Engineering Research Laboratory, International University of Business Agriculture and Technology, Dhaka-1230, Bangladesh"
}
],
"first_name": "Ferdousi",
"family_name": "Begum",
"email": null,
"author_order": 10,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 12
},
{
"id": 55,
"affiliation": [
{
"affiliation": "Chungbuk National University, Cheongju, Chungbuk 361-763, South Korea"
}
],
"first_name": "Probir Kumar",
"family_name": "Mittra",
"email": null,
"author_order": 11,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 12
},
{
"id": 56,
"affiliation": [
{
"affiliation": "Department of Agroforestry and Environmental Science, Sylhet Agricultural University, Sylhet 3100, Bangladesh"
}
],
"first_name": "Md. Omar",
"family_name": "Sharif",
"email": null,
"author_order": 12,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 12
},
{
"id": 57,
"affiliation": [
{
"affiliation": "College of Agricultural Sciences, International University of Business Agriculture and Technology, Dhaka-1230, Bangladesh"
}
],
"first_name": "Swapan Kumar",
"family_name": "Roy",
"email": "swapan.kumar@iubat.edu",
"author_order": 13,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": true,
"corresponding_author_info": "Swapan Kumar Roy, PhD; College of Agricultural Sciences, International University of Business Agriculture and Technology, Sector- 10, Dhaka-1230, Bangladesh. Email: swapan.kumar@iubat.edu",
"article": 12
}
],
"views": 1108,
"downloads": 243,
"references": [
{
"id": 349,
"serial_number": 1,
"reference": "Keddy PA. Plant ecology: Cambridge University Press; 2017.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 350,
"serial_number": 2,
"reference": "Wang SW, Boru BH, et al. Floristic composition and endemism pattern of vascular plants in Ethiopia and Eritrea. Journal of Systematics and Evolution. 2020;58:33-42.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 351,
"serial_number": 3,
"reference": "Huang H, Tu C, et al. Ecosystem complexity enhances the resilience of plant-pollinator systems. One Earth. 2021;4:1286-96.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 352,
"serial_number": 4,
"reference": "Agrawal A. Effect of global warming on climate change, flora and fauna. Journal of Ecophysiology and Occupational Health. 2011;11:161.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 353,
"serial_number": 5,
"reference": "Alam M, Pasha M. A floristic account of Chittagong university campus. Chittagong University Journal of Science. 1999;23:81-98.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 354,
"serial_number": 6,
"reference": "Sultana S, Uddin S, et al. A taxonomic study on the flora of Jahangirnagar university campus. Bangladesh J Pl Taxon. 2013.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 355,
"serial_number": 7,
"reference": "Mia M, Huq A. A preliminary ethnobotanical survey in the Jointiapur, Tamabil and Jafflong area, Sylhet, Bangladesh national herbarium bull. 1988.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 356,
"serial_number": 8,
"reference": "Haast B, O’Brien P. Population trends and issues. Education About Asia. 2019;24:66-70.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 357,
"serial_number": 9,
"reference": "Islam K, Sato N. Deforestation, land conversion and illegal logging in Bangladesh: The case of the sal (shorea robusta) forests. iForest-Biogeosciences and Forestry. 2012;5:171.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 358,
"serial_number": 10,
"reference": "Kumar R, Kumar A, et al. Deforestation and forests degradation impacts on the environment. Environmental degradation: Challenges and strategies for mitigation: Springer; 2022. p. 19-46.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 359,
"serial_number": 11,
"reference": "Naimuzzaman M, Rahman F, et al. Plant growth–promoting fungi in plants: Insights from stress tolerance mechanism. Beneficial microbes for sustainable agriculture under stress conditions: Elsevier; 2024. p. 469-511.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 360,
"serial_number": 12,
"reference": "Alemu B. The role of forest and soil carbon sequestrations on climate change mitigation. Res J Agr Environ Manage. 2014;3:492-505.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 361,
"serial_number": 13,
"reference": "Rahman MR. Causes of biodiversity depletion in Bangladesh and their consequences on ecosystem services. American journal of environmental protection. 2015;4:214-36.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 362,
"serial_number": 14,
"reference": "Roy S, Naimuzzaman M, et al. Glutathione: A key frontier of heavy-metal detoxification and toleranc e in plants. Plant Trends.1:33.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 363,
"serial_number": 15,
"reference": "Haque MSS, Rahman F, et al. Responses of nitrobenzene on growth and yield attributes in okra to salinity stress. Journal of Agricultural Sciences and Engineering. 2024;6:49-59.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 364,
"serial_number": 16,
"reference": "Khan TI, Islam MN, et al. Climate variability impacts on agricultural land use dynamics in the Madhupur tract in Bangladesh. Bangladesh I: Climate Change Impacts, Mitigation and Adaptation in Developing Countries. 2018:167-93.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 365,
"serial_number": 17,
"reference": "Islam M, Nusrat T, et al. Investigation of soil properties and nutrients in agricultural practiced land in tangail, Bangladesh. International Journal of Agricultural Research, Innovation and Technology (IJARIT). 2020;10:84-90.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 366,
"serial_number": 18,
"reference": "Lira S, Hossen M, et al. Quality assessment of Madhapur tract soil for agricultural activities. Bangladesh J. 2014;26:115-8.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 367,
"serial_number": 19,
"reference": "Prain D. Bengal plants. 1903.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 368,
"serial_number": 20,
"reference": "Ahmed Z, Begum Z, et al. Encyclopedia of flora and fauna of Bangladesh. 5-8 and 12, angiosperms: Dicotyledons and monocotyledons. Asiatic Society of Bangladesh, Dhaka; 2009.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 369,
"serial_number": 21,
"reference": "Ahmed Z, Tahmida Begum Z, et al. Encyclopedia of flora and fauna of Bangladesh. (No Title). 2008.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 370,
"serial_number": 22,
"reference": "Das D, Alam M. Trees of Bangladesh. Trees of Bangladesh. 2001.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 371,
"serial_number": 23,
"reference": "Ghani A. Medicinal plants of Bangladesh: Chemical constituents and uses: Asiatic society of Bangladesh; 1998.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 372,
"serial_number": 24,
"reference": "Khan MS, Rahman MM, et al. Red data book of vascular plants of Bangladesh. (No Title). 2001.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 373,
"serial_number": 25,
"reference": "Ljubojević M, Narandžić T, et al. Rethinking horticulture to meet sustainable development goals—the case study of Novi sad, Serbia. Horticulturae. 2022;8:1222.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 374,
"serial_number": 26,
"reference": "Agriculturels TU. Towards productive and socioмnatural urban landscapes: Tapping urban agriculturels potential as a tool for sustainable development.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 375,
"serial_number": 27,
"reference": "Shavanov M. The role of food crops within the Poaceae and Fabaceae families as nutritional plants. IOP Conference Series: Earth and Environmental Science: IOP Publishing; 2021. p. 012111.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 376,
"serial_number": 28,
"reference": "Werner D, Newton WE. Nitrogen fixation in agriculture, forestry, ecology, and the environment: Springer Science & Business Media; 2005.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 377,
"serial_number": 29,
"reference": "Vasconcelos MW, Grusak MA, et al. The biology of legumes and their agronomic, economic, and social impact. The plant family Fabaceae: biology and physiological responses to environmental stresses. 2020:3-25.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 378,
"serial_number": 30,
"reference": "de Haan S. Community-based conservation of crop genetic resources. Plant genetic resources: Burleigh Dodds Science Publishing; 2021. p. 229-49.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 379,
"serial_number": 31,
"reference": "Visioni A, Basile B, et al. Advancing the conservation and utilization of barley genetic resources: Insights into germplasm management and breeding for sustainable agriculture. Plants. 2023;12:3186.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 380,
"serial_number": 32,
"reference": "Planchuelo G, Kowarik I, et al. Plant traits, biotopes and urbanization dynamics explain the survival of endangered urban plant populations. Journal of Applied Ecology. 2020;57:1581-92.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 381,
"serial_number": 33,
"reference": "Aziz SA, McConkey KR, et al. The critical importance of old world fruit bats for healthy ecosystems and economies. Frontiers in Ecology and Evolution. 2021;9:641411.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 382,
"serial_number": 34,
"reference": "Phand SS, Ishwarya MY, et al. Agroforestry and opportunities in value addition.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 383,
"serial_number": 35,
"reference": "Hamza MA, Naimuzzaman M, et al. Health benefits of Moringa oleifera: Used as an anti-diabetic agent. International Journal of Agricultural Research, Innovation and Technology (IJARIT). 2023;13:96-102.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 384,
"serial_number": 36,
"reference": "Alagesan Venkatesan AV, Arumugam Kathirvel AK, et al. Antioxidant, antibacterial activities and identification of bioactive compounds from Terminalia chebula bark extracts. 2017.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 385,
"serial_number": 37,
"reference": "Pushpakumara G, Silva R, et al. Diversity of underutilized vegetables and fruit in Sri Lanka: Prioritization for collection, conservation, genetic improvement, and promotion. Working Paper. 2023.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 386,
"serial_number": 38,
"reference": "Sen TD. Domestication of some nutritionally important wild edible plants of Mandi Himachal pradesh, north western Himalayas for boosting health, immunity, rural prosperity and food security. 2021.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 387,
"serial_number": 39,
"reference": "Ogunniyi QA, Ogbole OO, et al. Medicinal importance and phytoconstituents of underutilized legumes from the Caesalpinioideae dc subfamily. Applied Sciences. 2023;13:8972.",
"pmc": null,
"DOI": null,
"article": 12
},
{
"id": 388,
"serial_number": 40,
"reference": "Warschefsky E. The evolution and domestication genetics of the mango genus, Mangifera (Anacardiaceae). 2018.",
"pmc": null,
"DOI": null,
"article": 12
}
]
},
{
"id": 11,
"slug": "280-1718510134",
"featured": true,
"slider": true,
"issue": "Vol2 Issue3",
"type": "original_article",
"manuscript_id": "280-1718510134",
"recieved": "2024-06-16",
"revised": null,
"accepted": "2024-07-12",
"published": "2024-07-27",
"pdf_file": "https://plant-trends.bsmiab.org/media/pdf_file/2024/05/280-1718510134.pdf",
"title": "An efficient approach of in vitro plant regeneration and propagation of mungbean [<span>Vigna radiata</span> L. (Wilczek)]",
"abstract": "<p>Mungbean [<em>Vigna radiata</em> L. (Wilczek)] is a valuable legume crop in Bangladesh. However, it faces severe difficulties forming viable roots during the growth stage. The successful root production of mungbean through an efficient tissue culture system has not been well established. Therefore, this study aims to investigate the problem of unsuccessful root formation in mungbean plants and to develop a very efficient method of <em>in vitro</em> regeneration using micropropagation. The BARI Mung-3 mungbean variety was used as an explant. Several shoots were produced from cotyledonary node (CN) explants obtained from 3-day-old seedlings that were germinated <em>in vitro</em>. The plant samples were cultivated on MSB5 medium enriched with 5.0 μM BAP. Shoot generation efficiency per plant was 5.36 ± 0.56 (80.08%). However, the viable root generation in a regulated setting was unsuccessful despite employing multiple combinations of rooting media, including full and half-strength MSB5 medium with different concentrations and combinations of auxins. To resolve this issue, a micrografting approach was applied with scion 3.0 cm in length and 14-day-old mungbean rootstocks. This system resulted in efficient shoots where the viable root generation efficiency rate was 55%. Interestingly, the micro-grafted plantlets successfully produced viable seeds. The successful micropropagation with viable root generation in mungbean plants successfully overcomes the difficulties in rooting and offers an efficient method for successful mungbean production. These new findings open new options for efficient plant generation with mass propagation for ready-smart mungbean production.</p>",
"journal_reference": "Plant Trends. 2024; 2(3): 46-56",
"academic_editor": "Md Mahadi Hasan, PhD; Lanzhou University, China",
"cite_info": "Bhajan SK, Hasan MM, et al. An efficient approach of in vitro plant regeneration and propagation of mungbean [Vigna radiata L. (Wilczek)]. Plant Trends. 2024; 2(3): 46-56.",
"keywords": [
"In vitro shoot regeneration",
"Micrografting",
"Rooting",
"Cotyledonary node",
"Legumes"
],
"DOI": "10.5455/pt.2024.05",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>Mungbean [<em>Vigna radiata</em> L. (Wilczek)],<em> </em>is commonly called green gram and is considered a highly significant pulse crop globally. This organism belongs to the Fabaceae family and the Papilionaceae subfamily. Legumes are regarded as the second most considerable food crop for humans. Legume seeds are a vital component of the human diet due to their high content of proteins, bioactive compounds, minerals, and vitamins. Compared to grains, legumes are often referred to as "the poor man's meat" [<a href=\"#r-1\">1</a>]. This legume crop is highly significant and widely cultivated, occupying over 6 million hectares globally (around 8.5% of the total pulse area). It is a staple in Asian families [<a href=\"#r-2\">2</a>]. The mung bean is rich in a well-balanced combination of nutrients, such as protein, dietary fiber, minerals, vitamins, and substantial quantities of bioactive substances [<a href=\"#r-3\">3</a>]. Mungbean grains are also widely popular for their high digestibility and good flavor. Mungbean sprouts are regarded as a vegetable high in vitamin C and contain 12 times as much iron as regular Mungbean seeds [<a href=\"#r-4\">4</a>]. Nevertheless, the productivity of Mungbean varieties grown in various regions worldwide varies, and their potential yields are known to be influenced by factors such as environmental conditions, pests and diseases, inadequate farming techniques, and limited access to high-quality seeds of new varieties for farmers [<a href=\"#r-5\">5</a>]. The main factor contributing to reduced crop yield is its vulnerability to several diseases, including yellow mosaic virus, <em>Cercospora</em> leaf spot, powdery mildew, and root rot [<a href=\"#r-6\">6</a>, <a href=\"#r-7\">7</a>]. The Mungbean yellow mosaic virus (MYMV) is regarded as the most destructive disease of Mungbean in South Asian countries, especially Bangladesh [<a href=\"#r-8\">8</a>, <a href=\"#r-9\">9</a>]. Conventional breeding produced limited success in obtaining virus disease resistance in Mungbean. Thus, it is hypothesized that the enhancement of this significant crop can be accomplished by employing advanced biotechnology techniques, such as plant genetic engineering. It is widely acknowledged that the successful implementation of plant genetic transformation requires an effective <em>in vitro</em> plant regeneration system. Mungbean is recalcitrant and the number of regenerating shoots remained very low and limited to a few only, That’s why suitable tissue culture methods are needed for the improvement of Mungbean through micropropagation [<a href=\"#r-10\">10</a>].</p>\r\n\r\n<p>In the past, several attempts have been made to establish a suitable, effective reproducible <em>in vitro </em>regeneration protocol for Mungbean using different explants [<a href=\"#r-11\">11-16</a>]. Developing such regeneration has been reported to be more difficult in mungbean than that obtained in other grain legumes [<a href=\"#r-14\">14</a>]. Notably, recovery of the regenerated plants significantly depends on the development of viable roots in vitro. The subsequent recovery of the transformed shoots has always been a great hurdle due to the recalcitrant nature of mungbean explants towards <em>in vitro</em> root generation. This ultimately greatly reduces the transformation efficiency in mungbean [<a href=\"#r-17\">17</a>], making the process expensive, labour-intensive, and time-consuming, and ultimately hindering the widespread adoption of genetic modification in mungbean as a method of genetic improvement. The establishment of robust root systems in potential transformants is sometimes challenging due to extended exposure to high antibiotic concentrations throughout the selection process. Mungbean transformants maintained under prolonged antibiotic treatment yielded the same outcomes in a different set of experiments. In these conditions, micrografting might be a solution to the issue of successful root formation from <em>in vitro</em>-grown shoots of mungbean.</p>\r\n\r\n<p>Micrografting is performed by grafting a meristem tip, a shoot tip, or a newly developed shoot onto a decapitated rootstock, aseptically. This technique has several benefits, including the eradication of viruses, year-round plant production, the creation of specialized genotypic combinations to boost plant productivity, and the development of genotypic combinations to augment plant yield and extend the ecological limits of a particular plant species [<a href=\"#r-18\">18</a>]. The investigation of the histological characteristics of graft unions has also been made easier by micrografting [<a href=\"#r-19\">19</a>]. The micrografting approach has not been used in root generation in mungbean. Therefore, successful regeneration and micropropagation of mungbean would be a promising and efficient approach.</p>\r\n\r\n<p>To develop <em>in vitro</em> micro propagated plants, the problems addressed with <em>in vitro</em> root production and micrografting technique followed the possible solution [<a href=\"#r-20\">20-22</a>]. Successful micrografting has been observed in various plants, such as cotton, fruit trees, species of woody plants, and a Leguminosae family member closely related to lentils and chickpeas [<a href=\"#r-23\">23-26</a>]. There are no such reports discussing the use of micrografting to address issues with the growth of full plantlets in the case of Mungbean cultivars. Therefore, to successfully recover plants from <em>in vitro</em> regenerated shoots the techniques of micrografting were utilized. Thus, this study aimed to create an effective grafting method for Mungbean to help <em>in vitro</em> regeneration of complete plants.</p>"
},
{
"section_number": 2,
"section_title": "MATERIALS AND METHODS",
"body": "<p><strong>Plant seed collection and processing </strong></p>\r\n\r\n<p>The present study utilized mungbean [<em>Vigna radiata</em> L. (Wilczek)] seeds<em> </em>of the BARI Mung-3 variety, sourced from the Bangladesh Agricultural Research Institute (BARI) at Joydebpur, Gazipur. Cotyledonary node (CN) explants were obtained from 3-day-old seedlings grown <em>in vitro</em>. The explants were generated by detaching 4.0 mm sections of both the epicotyl and hypocotyl.</p>\r\n\r\n<p><strong> </strong></p>\r\n\r\n<p><strong>Media and growth condition</strong></p>\r\n\r\n<p>For <em>in-vitro</em> regeneration, MSB5 media BAP and IBA hormone were used in this present study. The composition of the MSB5 medium is followed by Gamborg [<a href=\"#r-27\">27</a>]. The cultures were maintained at a temperature of 25 ±1°C with a 16/8-hour (light/dark) photoperiod using a cool white, fluorescent light (3000 lux). A fluorescent microscope was utilized to capture an image of the process of graft union development.</p>\r\n\r\n<p><strong><em> </em></strong></p>\r\n\r\n<p><strong><em>In vitro</em></strong><strong> micropropagation</strong></p>\r\n\r\n<p>To prepare CN explants, we sterilized healthy and uniform seeds by rinsing them with tap water and then washing them with 70% ethanol for 1 minute. To enhance the sterility of the seeds, a solution of 0.1% HgCl<sub>2</sub> (W/V) was applied for 10 minutes. This was followed by 4-5 cycles of rinsing with sterilized distilled water. The seeds that had been sterilized on the surface were germinated in a sterile manner on Petri plates containing half-strength MS medium and 0.8% agar. The germination process took place at a temperature of 25°C under dark conditions [<a href=\"#r-28\">28</a>]. To induce shoot regeneration, the plant tissue samples were placed on MSB5 medium supplemented with different concentrations of BAP (0.0, 2.5, 5.0, 7.5, and 10.0 μM) [<a href=\"#r-29\">29</a>]. In addition, the explants were primarily cultivated for 30 days, with sub-culturing occurring at intervals of 15 days on the medium indicated earlier.</p>\r\n\r\n<p><strong> </strong></p>\r\n\r\n<p><strong>Micrografting</strong></p>\r\n\r\n<p>Once the shoots had undergone enough growth and elongation, they were transferred to a rooting medium with half the strength of MSB5 medium. This medium was treated with different concentrations (1.0, 1.5, 2.0, and 2.5 μM) of IBA. For micrografting, scions consisting of shoots that were produced in vitro and were 28 days old were utilized. The rootstocks were created using seedlings that were grown in vitro for either 7 or 14 days. To prepare the rootstocks, the shoot tip, axillary buds, and cotyledons were removed from the seedlings using a surgical blade. Upon removing the upper sections, the uppermost cut surface of the shoot was longitudinally split open to a depth of 50 mm. To achieve these objectives shoots measuring 1.5 - 3.0 cm in length were carefully removed as scions. A cut in the shape of a "V" was then made at the base of the scion. The grafting process involved gently inserting the scion into the vertical split of the rootstock, aligning a portion of the cambium layers of both the scion and rootstock (<a href=\"#figure1\">Figure 1</a>). Following the insertion of the scion, the grafted area was securely sealed with masking tape and then enveloped in a translucent polythene bag to ensure optimal humidity levels. The process of strengthening micrografted plantlets was carried out at optimal room temperature conditions at 25±1°C, with a day length of 16 hours, until the graft union was successfully formed. During the initial 14-day period, the bags were systematically punctured to facilitate the exchange of air for the purpose of plant acclimation.</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"244\" src=\"/media/article_images/2024/34/28/280-1718510134-figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1.</strong> Diagram illustrating the procedure of micrografting. Scions are derived from shoots that have been regenerated in vitro and then sliced into 'V' shapes. A seedling that has been grown in a laboratory setting and had its top part removed at the point where the stem emerges is shown to have the rootstock cut in a similar 'V' shape. The process of grafting shoot tips onto the rootstock is achieved through top grafting using masking tape. A micrografted plant has been successfully produced, and the circular marking marks the specific place where the micrograft has been performed.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Transplantation</strong></p>\r\n\r\n<p>Following the process of acclimatization, the plants that were successfully grafted were relocated to larger earthen pots within a net house, where they were exposed to a natural environment until the growth of flowers and pods. The survival rate of the grafted plants was assessed after 30 to 40 days by analyzing the data that had been collected. Moreover, anatomical studies were done to comprehend how graft union was established in the case of Mungbean by using a fluorescent microscope.</p>\r\n\r\n<p><strong> </strong></p>\r\n\r\n<p><strong>Statistical analysis</strong></p>\r\n\r\n<p>The experiment was repeated three times, with each treatment being replicated 20 times. The studies were conducted in the laboratory using a completely randomized design (CRD). The data were statistically examined using Duncan's multiple-range test. The data obtained from several metrics, such as the percentage of regeneration, the number of shoots per explant, and the percentage of root initiation, were computed using different statistical measures: percentage, average, and standard deviation, respectively.</p>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p><strong>Successful shoot regeneration of mungbean variety BARI Mung-3</strong></p>\r\n\r\n<p>Several tests were conducted using aseptically grown explants (CN) of the Mungbean variety BARI Mung-3 to accomplish shoot regeneration in vitro. In order to achieve this objective, we used MSB5 media supplemented with varying amounts of BAP and kinetin to induce the growth of shoots in vitro. We utilized cotyledonary node explants for this purpose, as shown in <a href=\"#figure2\">Figure 2</a>a. The impact of varying quantities of BAP and kinetin on the process of shoot regeneration in the Mungbean variety was observed to be inconsistent, as indicated in <a href=\"#Table-1\">Table 1</a>. Upon the addition of 5.0 M BAP to MSB5 medium, almost 80% of cotyledonary node explants exhibited a positive response to shoot induction. The BARI Mung-3 variety had the largest average number of shoots per explant, which was 5.36 ± 0.56 (80.08%) (<a href=\"#Table-1\">Table 1</a>).</p>\r\n\r\n<div id=\"Table-1\">\r\n<p><a href=\"https://plant-trends.bsmiab.org/table/280-1718510134-table1/\">Table-1</a><strong>Table 1. </strong>Investigation of the effects of varying doses of BAP and their kinetics on shoot regeneration from cotyledonary node explants of the BARI Mung 3 variety.</p>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Production of efficient rooting of mungbean </strong></p>\r\n\r\n<p>It was recorded that on average 12 days were required for the initiation of shoot from cotyledonary node explants (<a href=\"#figure2\">Figure 2</a>b), while the formation of multiple shoots occurred at least 30 days from the initiation of culture. Following this experiment,<em> in vitro</em> raised multiple shoots have been presented in <a href=\"#figure2\">Figure 2</a>c. After the development of sufficient healthy shoots, they were cultured in different media for the induction of roots. To do this, 3-5 cm long shoots were cut off and cultured on half-strength MSB media with different IBA concentrations (<a href=\"#Table-2\">Table 2</a>). Viable and functional root development was not successful due to the small number of roots that emerged and were not elongated enough to facilitate successful transplantation in the soil. Besides, the survival rate of the <em>in vitro</em> rooted plantlets was extremely low. So, in this case, the overall performance of the <em>in vitro</em> root development was not satisfactory. Under these circumstances, micrografting techniques were employed to get around obstacles to proper root formation and to increase the survival of <em>in vitro</em> generated plantlets.</p>\r\n\r\n<div id=\"Table-2\">\r\n<p><a href=\"https://plant-trends.bsmiab.org/table/280-1718510134-table2/\">Table-2</a><strong>Table 2. </strong>The impact of varying concentrations of IBA on the development of roots from in vitro propagated shoots in the BARI mung-3 variety was investigated using a half-strength MSB<sub>5</sub> medium.</p>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"332\" src=\"/media/article_images/2024/34/28/280-1718510134-figure2.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 2</strong>. <em>In vitro </em>regeneration of shoots and stages of micrografting in BARI Mung 3 variety. a) Cotyledonary node (CN) explants excised from 3-day-old <em>in vitro </em>grown seedlings. b) Initiation of <em>in vitro </em>shoots from CN explant. c) Multiple shoots developing from CN explant on MSB<sub>5</sub> medium supplemented with 5.0 μM BAP. d) <em>In vitro, </em>raised shoots are used as scions for grafting. e) Seedlings prepared to use as rootstock for micrografting, and f) Successful micrografted plants after 33 days of grafting. Arrows indicate the grafted region covered by masking tape. </figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Scion length and rootstock age are vital factors for successful micrografting </strong></p>\r\n\r\n<p>Several factors were found to be associated with the development of a protocol for micrografting in Mungbean. It was recorded that scions (Figure 2d) having a length of 2.5 - 3.0 cm exhibited better response in establishing micrografts with a success rate of 55 % (<a href=\"#Table-3\">Table 3</a>). The second highest rate of grafting (48%) was obtained from the scions, which had a length of 1.5–2.0 cm. Identical observations regarding the role of scion length in graft formation were reported in the case of sandalwood plants [<a href=\"#r-30\">30</a>]. It was observed that the 14-day-old rootstocks (<a href=\"#figure2\">Figure 2</a>e) were more efficient than the 7-day-old rootstocks in achieving the desired micrografts (<a href=\"#Table-3\">Table 3</a>).</p>\r\n\r\n<div id=\"Table-3\">\r\n<p><a href=\"https://plant-trends.bsmiab.org/table/280-1718510134-table3/\">Table-3</a><strong>Table 3. </strong>The impact of scion length and rootstock age on the success rate of grafting.</p>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Factors associated with decreased rate or failure of micrografting </strong></p>\r\n\r\n<p>The success rate of grafting decreased when the age of the rootstock exceeded 28 days. It was also noticed that the survival of the successful grafts was unaffected by the scion insertion and the extraction of the cotyledons from the rootstock. Despite the grafting method being less complicated and easy to perform, the chances of failure in grafting (63%) with the young rootstocks (7 days old) and smaller scions (1.5 - 2.0 cm) lengths. The displacement of the microscope has failed micrografting. The average success rate of grafting in mungbean plants was 51% (<a href=\"#figure2\">Figure 2</a>f). After the establishment of a successful graft, the average success rate of transplantation was 81.66 (<a href=\"#Table-4\">Table 4</a>). The plantlets that were propagated grew very well and exhibited no morphological abnormalities. Consequently, Mungbean plants that had undergone micrografting had normal flowering and pod development (Figures 3a and 3b). The pods reached maturity and bore viable seeds (<a href=\"#figure3\">Figure 3</a>c).</p>\r\n\r\n<p>To comprehend the nature of graft union anatomical studies were carried out particularly several transverse sections were done through the regions of graft formation and were observed under the microscope. To develop graft union formation between rootstock and scion the most</p>\r\n\r\n<p>An important advancement is the establishment of vascular continuity across the interface zone. Successful grafting occurred only when there was a complete fusion of the xylem and phloem of the scion and rootstock. The integration of vascular bundles between the scion and rootstock was observed to be fully established 30 days following the grafting process. Callus production is a crucial phase for the establishment of a vascular bundle since it effectively connects the scion to the rootstock. Based on current observations, cellular division initiates from the canter region of the union where the rootstock and scion are united. Various stages of graft union are presented in <a href=\"#figure4\">Figure 4</a>a. A fluorescent micrograph of the transverse section of the stem showing the position of vascular tissue (<a href=\"#figure4\">Figure 4</a>b). Fluorescent microscopic observation through the transverse section demonstrated the position of the vascular tissues of Mungbean shoot as well as the consequences graft union particularly the changes that occurred within the xylem and phloem. These observations indicated the establishment of the connectivity between the vascular tissues of the scion and rootstock (<a href=\"#figure4\">Figure 4</a>c).</p>\r\n\r\n<div id=\"figure3\">\r\n<figure class=\"image\"><img alt=\"\" height=\"238\" src=\"/media/article_images/2024/34/28/280-1718510134-figure3.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 3</strong>. Development of flowers and pods on micrografted plants of BARI Mung 3 variety. a) Flower developed on a grafted plant indicating the successful establishment of a grafted plant. b) Pod formation on micrografted plants. c) Several healthy seeds were obtained from pods developed on the micrografted plant of Mungbean.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"figure4\">\r\n<figure class=\"image\"><img alt=\"\" height=\"206\" src=\"/media/article_images/2024/34/28/280-1718510134-figure4.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 4</strong>. Graft union and anatomy of graft formation. a) Various stages of graft union, i. a control shoot, ii. & iii showing stages of establishment of graft, iv. fully establish a micrografted region. Arrows indicate a developing grafted region. b) A fluorescent micrograph of a transverse section of a control shoot showing the position of vascular tissues (arrows). c) A fluorescent micrograph shows the connected vascular tissues (arrows) of the scion (sc) and rootstock (rs).</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"Table-4\">\r\n<p><a href=\"https://plant-trends.bsmiab.org/table/280-1718510134-table4/\">Table-4</a><strong>Table 4. </strong>The postoperative success rate of grafts and transplants following 30 days.</p>\r\n</div>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p>Mungbean is a recalcitrant plant, presenting significant challenges in developing complete plantlets through micropropagation. One of the primary difficulties in micropropagation for some species is inducing adventitious root formation [<a href=\"#r-31\">31</a>]. Under those circumstances, micrografting is an alternate method used in vitro to facilitate root growth and overcome challenges in rooting during the vegetative development of these species. Numerous genotypes of leguminous crop plants have reported experiencing such challenges in developing functional roots [<a href=\"#r-32\">32</a>].<strong> </strong>More particularly, several investigations have described issues with the development of <em>in vitro</em> roots from regenerated shoots in grain legumes like lentils, peas, and chickpeas [<a href=\"#r-24\">24</a>, <a href=\"#r-33\">33-36</a>] as difficult. Additionally, it has been documented that in certain leguminous species, the prolonged presence of auxin in the rooting medium did not facilitate the typical rooting process from regenerated shoots [<a href=\"#r-11\">11</a>]. While the use of full-strength MS media is common for in vitro rooting in various species, the high quantities of macronutrients in this medium can impede root initiation and lead to shoot necrosis after prolonged incubation [<a href=\"#r-37\">37</a>, <a href=\"#r-38\">38</a>]. In the case of transgenic shoots, the <em>in vitro</em> regenerated shoots are cultured for a long period under antibiotic selection conditions and different BAP hormones that create a problem for proper root development. There are various reports that <em>in vitro</em> rooting in chickpeas has a negative correlation with added BAP concentration resulting in a 35% decrease in rooting [<a href=\"#r-39\">39</a>, <a href=\"#r-40\">40</a>]. BAP negatively affected the <em>in vitro</em> rooting of Mungbean during this study (data not shown), and many other crops have shown comparable effects. Moreover, in many instances when phenolic compounds are released, the growth of tissue in culture stops, which prevents root initiation and development and ultimately causes explant mortality. These phenolics easily oxidize, which results in tissue blackening and medium browning [<a href=\"#r-41\">41</a>].<strong> </strong>So micrografting of Mungbean is a good alternative to produce a viable complete plant through micropropagation.</p>\r\n\r\n<p>Various factors affect this micro grafting technique. It was found that the length of the scion and the age of the rootstock employed impacted the rate of successfully grafted shoots during the current micrografting experiments. According to Nelson 2006 [<a href=\"#r-42\">42</a>], in the case of <em>Acacia</em><em> koa, </em>the grafting<em> </em>fails due to the narrow stem diameter of the rootstock and young stem tissue of the scion used for ensuring proper positioning of the microscion onto the rootstock is crucial for establishing adequate contact, which is important for the formation of the graft union. The success rate of rooting of normal regenerated shoots is 26.66 or lower when using selection pressure but after <em>in vitro</em> micrografting the success rate is more than 55%. The development of vascular cambium tissues of the rootstock and scion plants must be placed in contact with each other for a successful grafting to take place, in my study good vascular connectivity occurs in mungbean plants. Prior studies have documented the successful micrografting of shoots in several grain legumes and fruit crops e.g. citrus [<a href=\"#r-26\">26</a>], lentil [<a href=\"#r-24\">24</a>], <em>Pisum sativum</em> [<a href=\"#r-34\">34</a>], chickpea [<a href=\"#r-35\">35</a>], horticultural crops [<a href=\"#r-18\">18</a>], and woody plant species [<a href=\"#r-43\">43</a>].</p>"
},
{
"section_number": 5,
"section_title": "CONCLUSION",
"body": "<p>The present findings suggest that micrografting offers a robust solution to the rooting challenges in mungbean plants with successful root regeneration, providing a practical approach for plant tissue culture with vegetative propagation-based trait improvement projects. This study suggests the efficient plant tissue culture approach as well as the efficient plant regeneration technique of mungbean plants. This study could be useful to legume breeders or farmers for improving mungbean and other legume species using breeding programs.</p>"
},
{
"section_number": 6,
"section_title": "AUTHOR CONTRIBUTIONS",
"body": "<p>SKB conducted the investigation, wrote the original manuscript, and created the figures. MMH conducted formal analysis and data generation, with references included. Revisions and evaluation by MAR. Conceptualization, draft editing, modification, and supervision were performed by RHS and MNI. All authors have reviewed and consented to the final version of the manuscript that has been published.</p>"
},
{
"section_number": 7,
"section_title": "ACKNOWLEDGMENTS",
"body": "<p>The authors are grateful for the financial support provided by the BAS-USDA research grant. They would also like to thank BARI for providing mungbean seeds used in this study. I am deeply grateful to my late Ph.D. supervisor, Professor Dr. M. Imdadul Hoque, Department of Botany, University of Dhaka, for his invaluable guidance, suggestions, and encouragement, which made the research project successful.<strong> </strong></p>"
},
{
"section_number": 8,
"section_title": "CONFLICTS OF INTEREST",
"body": "<p>There is no conflict of interest among the authors.</p>"
}
],
"figures": [
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/34/28/280-1718510134-figure1.jpg",
"caption": "Figure 1. Diagram illustrating the procedure of micrografting. Scions are derived from shoots that have been regenerated in vitro and then sliced into 'V' shapes. A seedling that has been grown in a laboratory setting and had its top part removed at the point where the stem emerges is shown to have the rootstock cut in a similar 'V' shape. The process of grafting shoot tips onto the rootstock is achieved through top grafting using masking tape. A micrografted plant has been successfully produced, and the circular marking marks the specific place where the micrograft has been performed.",
"featured": false
},
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/34/28/280-1718510134-figure2.jpg",
"caption": "Figure 2. In vitro regeneration of shoots and stages of micrografting in BARI Mung 3 variety. a) Cotyledonary node (CN) explants excised from 3-day-old in vitro grown seedlings. b) Initiation of in vitro shoots from CN explant. c) Multiple shoots developing from CN explant on MSB5 medium supplemented with 5.0 μM BAP. d) In vitro, raised shoots are used as scions for grafting. e) Seedlings prepared to use as rootstock for micrografting, and f) Successful micrografted plants after 33 days of grafting. Arrows indicate the grafted region covered by masking tape.",
"featured": true
},
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/34/28/280-1718510134-figure3.jpg",
"caption": "Figure 3. Development of flowers and pods on micrografted plants of BARI Mung 3 variety. a) Flower developed on a grafted plant indicating the successful establishment of a grafted plant. b) Pod formation on micrografted plants. c) Several healthy seeds were obtained from pods developed on the micrografted plant of Mungbean.",
"featured": false
},
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/34/28/280-1718510134-figure4.jpg",
"caption": "Figure 4. Graft union and anatomy of graft formation. a) Various stages of graft union, i. a control shoot, ii. & iii showing stages of establishment of graft, iv. fully establish a micrografted region. Arrows indicate a developing grafted region. b) A fluorescent micrograph of a transverse section of a control shoot showing the position of vascular tissues (arrows). c) A fluorescent micrograph shows the connected vascular tissues (arrows) of the scion (sc) and rootstock (rs).",
"featured": false
}
],
"authors": [
{
"id": 40,
"affiliation": [
{
"affiliation": "Department of Biotechnology and Genetic Engineering, Bangabandhu Sheikh Mujibur Science and Technology University, Gopalganj, Bangladesh"
}
],
"first_name": "Sujay Kumar",
"family_name": "Bhajan",
"email": null,
"author_order": 1,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 11
},
{
"id": 41,
"affiliation": [
{
"affiliation": "Department of Microbiology and Cell Science, University of Florida, Florida, USA"
}
],
"first_name": "Mohammad Mahmood",
"family_name": "Hasan",
"email": null,
"author_order": 2,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 11
},
{
"id": 42,
"affiliation": [
{
"affiliation": "Global Biotechnology & Biomedical Research Network (GBBRN), Department of Biotechnology and Genetic Engineering, Faculty of Biological Sciences, Islamic University, Kushtia-7003, Bangladesh"
}
],
"first_name": "Md Ataur",
"family_name": "Rahman",
"email": null,
"author_order": 3,
"ORCID": "https://orcid.org/0000-0001-6649-3694",
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 11
},
{
"id": 43,
"affiliation": [
{
"affiliation": "Plant Breeding and Biotechnology Laboratory, Department of Botany, University of Dhaka, Dhaka-1000, Bangladesh"
}
],
"first_name": "Rakha Hari",
"family_name": "Sarker",
"email": null,
"author_order": 4,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 11
},
{
"id": 44,
"affiliation": [
{
"affiliation": "Plant Breeding and Biotechnology Laboratory, Department of Botany, University of Dhaka, Dhaka-1000, Bangladesh"
}
],
"first_name": "Mohammad Nurul",
"family_name": "Islam",
"email": "mnurul@du.ac.bd",
"author_order": 5,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": true,
"corresponding_author_info": "Mohammad Nurul Islam, PhD ; Plant Breeding and Biotechnology Laboratory, Department of Botany, University of Dhaka, Dhaka-1000, Bangladesh. Email: mnurul@du.ac.bd",
"article": 11
}
],
"views": 1040,
"downloads": 234,
"references": [
{
"id": 306,
"serial_number": 1,
"reference": "Hall C, Hillen C, et al. Composition, nutritional value, and health benefits of pulses. Cereal Chemistry. 2017;94:11-31.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 307,
"serial_number": 2,
"reference": "Hou D, Yousaf L, et al. Mung bean (vigna radiata l.): Bioactive polyphenols, polysaccharides, peptides, and health benefits. Nutrients. 2019;11:1238.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 308,
"serial_number": 3,
"reference": "Dahiya P, Linnemann A, et al. Mung bean: Technological and nutritional potential. Critical reviews in food science and nutrition. 2015;55:670-88.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 309,
"serial_number": 4,
"reference": "Kumar S, Pandey G. Biofortification of pulses and legumes to enhance nutrition. Heliyon 6 (3): E03682. 2020.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 310,
"serial_number": 5,
"reference": "Pratap A, Gupta S, et al. Towards development of climate smart mungbean: Challenges and opportunities. 2019:235-64.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 311,
"serial_number": 6,
"reference": "HanumanthaRao B, Nair RM, et al. Salinity and high temperature tolerance in mungbean [Vigna radiata (L.) Wilczek] from a physiological perspective. 2016;7:186318.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 312,
"serial_number": 7,
"reference": "Mishra GP, Dikshit HK, et al. Yellow mosaic disease (ymd) of mungbean (Vigna radiata (L) Wilczek): Current status and management opportunities. 2020;11:918.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 313,
"serial_number": 8,
"reference": "Nariani TJIP. Yellow mosaic of mung (Phaseolus aureus L.). 1960;13.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 314,
"serial_number": 9,
"reference": "Jalaluddin M, Shaikh M. Evaluation of mungbean (Vigna radiata (L.) Willczek) germplasm for resistance to mungbean yellow mosaic virus. 1981.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 315,
"serial_number": 10,
"reference": "Yadav S, Sreenu P, et al. Efficient shoot regeneration from double cotyledonary node explants of green gram [vigna radiata (l.) wilczek]. 2010.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 316,
"serial_number": 11,
"reference": "Gulati A, Jaiwal PKJPCR. Plant regeneration from cotyledonary node explants of mungbean (Vigna radiata (L.) Wilczek). 1994;13:523-7.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 317,
"serial_number": 12,
"reference": "Chandra M, Pal AJPCR. Differential response of the two cotyledons of Vigna radiata in vitro. 1995;15:248-53.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 318,
"serial_number": 13,
"reference": "Amutha S, Ganapathi A, et al. In vitro organogenesis and plant formation in Vigna radiata (L.) Wilczek. 2003;72:203-7.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 319,
"serial_number": 14,
"reference": "Khatun M, Haque M, et al. In vitro regeneration of mungbean (Vigna radiata L.) from different explants. 2008;19:13-9.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 320,
"serial_number": 15,
"reference": "Yadav S, Maheswari M, et al. Efficient shoot regeneration from double cytoledonary node explants of greengram [Vigna radiata (L.) Wilczek]. 2010.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 321,
"serial_number": 16,
"reference": "Bhajan SK, Begum S, et al. In vitro regeneration and agrobacterium-mediated genetic transformation of local varieties of mungbean (Vigna radiata (L). Wilczek). 2019:81-97.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 322,
"serial_number": 17,
"reference": "Sanjaya, Muthan B, et al. Micropropagation of an endangered indian sandalwood (Santalum album L.). 2006;11:203-9.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 323,
"serial_number": 18,
"reference": "Hussain G, Wani M, et al. Micrografting for fruit crop improvement. 2014;13.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 324,
"serial_number": 19,
"reference": "Abousalim A, Mantell SJPc, tissue, et al. Micrografting of pistachio (Pistacia vera L. Cv. Mateur). 1992;29:231-4.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 325,
"serial_number": 20,
"reference": "Sanyal I, Singh AK, et al. Agrobacterium-mediated transformation of chickpea (Ccer arietinum L.) with bacillus thuringiensis cry1ac gene for resistance against pod borer insect helicoverpa armigera. 2005;168:1135-46.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 326,
"serial_number": 21,
"reference": "Wang G, Li X, et al. Studies on factors affecting the microshoot grafting survival of walnut. VI International Walnut Symposium 8612009. p. 327-32.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 327,
"serial_number": 22,
"reference": "Yıldırım H, Akdemir H, et al. In vitro micrografting of the almond cultivars “texas”,“ferrastar” and “nonpareil”. 2013;27:3493-501.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 328,
"serial_number": 23,
"reference": "Banerjee A, Agrawal D, et al. Recovery of in vitro cotton shoots through micrografting. 2000:623-6.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 329,
"serial_number": 24,
"reference": "Gulati A, Schryer P, et al. Regeneration and micrografting of lentil shoots. 2001;37:798-802.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 330,
"serial_number": 25,
"reference": "Tsutsui H, Notaguchi MJP, et al. The use of grafting to study systemic signaling in plants. 2017;58:1291-301.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 331,
"serial_number": 26,
"reference": "Singh AK, Meetei NT, et al. In vitro micrografting using three diverse indigenous rootstocks for the production of citrus tristeza virus-free plants of khasi mandarin. 2019;55:180-9.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 332,
"serial_number": 27,
"reference": "Gamborg OLc, Miller RA, et al. Nutrient requirements of suspension cultures of soybean root cells. Experimental cell research. 1968;50:151-8.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 333,
"serial_number": 28,
"reference": "Murashige T, Skoog FJPp. A revised medium for rapid growth and bio assays with tobacco tissue cultures. 1962;15.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 334,
"serial_number": 29,
"reference": "Patra AP, Samal KC, et al. In vitro regeneration of recalcitrant green gram [Vigna radiata (L.) Wilczek] from immature cotyledons for genetic improvement. 2018;7:3072-80.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 335,
"serial_number": 30,
"reference": "Sanjaya, Muthan B, et al. Micropropagation of an endangered indian sandalwood (Santalum album L.). Journal of Forest Research. 2006;11:203-9.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 336,
"serial_number": 31,
"reference": "Wang M-R, Bettoni JC, et al. In vitro micrografting of horticultural plants: Method development and the use for micropropagation. 2022;8:576.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 337,
"serial_number": 32,
"reference": "Dewir YH, Murthy HN, et al. In vitro rooting of leguminous plants: Difficulties, alternatives, and strategies for improvement. 2016;57:311-22.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 338,
"serial_number": 33,
"reference": "Williams DJ, McHughen AJPc, tissue, et al. Plant regeneration of the legume Lens culinaris Medik.(lentil) in vitro. 1986;7:149-53.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 339,
"serial_number": 34,
"reference": "Mohommad Arif MA, Singh R, et al. Micrografting of in vitro cultured shoots of Pisum sativum L. Onto potted root stocks. 2009.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 340,
"serial_number": 35,
"reference": "Yadav IS, Singh NJJofl. In vitro regeneration and micrografting of shoots for enhancing survival and recovery of plants in chickpea (Cicer arietinum L.). 2012;25:97-101.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 341,
"serial_number": 36,
"reference": "Sarker R, Das SK, et al. In vitro flowering and seed formation in lentil (Lens culinaris Medik.). 2012;48:446-52.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 342,
"serial_number": 37,
"reference": "Parveen S, Shahzad AJP, et al. Tdz-induced high frequency shoot regeneration in cassia sophera linn. Via cotyledonary node explants. 2010;16:201-6.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 343,
"serial_number": 38,
"reference": "Agrawal V, Sardar PRJIVC, et al. In vitro regeneration through somatic embryogenesis and organogenesis using cotyledons of cassia angustifolia vahl. 2007;43:585-92.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 344,
"serial_number": 39,
"reference": "Fernandez-Romero M, Moreno M, et al. An efficient in vitro rooting method for chickpea. 1998.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 345,
"serial_number": 40,
"reference": "Polisetty R, Paul V, et al. Multiple shoot induction by benzyladenine and complete plant regeneration from seed explants of chickpea (Cicer arietinum L.). 1997;16:565-71.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 346,
"serial_number": 41,
"reference": "Ahmad I, Hussain T, et al. Lethal effects of secondary metabolites on plant tissue culture. 2013;13:539-47.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 347,
"serial_number": 42,
"reference": "Nelson SCJNpj. Grafting of acacia koa gray onto young acacia seedlings. 2006;7:137-40.",
"pmc": null,
"DOI": null,
"article": 11
},
{
"id": 348,
"serial_number": 43,
"reference": "Chandran S, Sarangapani S, et al. Micrografting: A in-vitro technique for sustainable horticulture production and its applications. 2023.",
"pmc": null,
"DOI": null,
"article": 11
}
]
},
{
"id": 10,
"slug": "280-1715253508",
"featured": false,
"slider": false,
"issue": "Vol2 Issue2",
"type": "original_article",
"manuscript_id": "280-1715253508",
"recieved": "2024-05-09",
"revised": null,
"accepted": "2024-06-16",
"published": "2024-06-29",
"pdf_file": "https://plant-trends.bsmiab.org/media/pdf_file/2024/16/280-1715253508.pdf",
"title": "In vitro evaluation of antioxidant and antimicrobial properties of <span>Cordia mocleodii</span> leaf extract",
"abstract": "<p><em>Cordia macleodii</em> is known as a medicinal plant with hepatoprotective and wound-healing properties. <em>Cordia macleodii</em> contains relatively high levels of flavonoids, alkaloids, steroids, and terpenoids. This study aimed to investigate the <em>in vitro</em> antibacterial and antioxidant properties of <em>Cordia macleodii</em> methanol leaf extract. The antioxidant activity of the methanol extract of <em>Cordia macleodii</em> leaves was determined using the DPPH scavenging test, total phenol content (TPC), and total flavonoid content (TFC). The antibacterial activity was evaluated using the disc diffusion method. The extract exhibited significant dose-dependent antioxidant activity comparable to ascorbic acid. The total flavonoid content was 611.9 mg/g, and the total phenol content was 164.4 mg/g. The DPPH free radical scavenging assay indicated an ascorbic acid production of 523.21 μg/ml, while routine tests showed 18.35 μg/ml. Additionally, the leaf extracts demonstrated strong antibacterial activity against various bacteria, even exceeding the effectiveness of Ciprofloxacin. Further research is necessary to develop targeted therapies, potentially opening new avenues for harnessing the medicinal properties of <em>Cordia macleodii</em>. Clinical studies are also needed to investigate its potential as an alternative medicine.</p>",
"journal_reference": "Plant Trends. 2024; 2(2): 38-45",
"academic_editor": "Md Jamal Uddin, PhD; ABEx Bio-Research Center, Dhaka, Bangladesh",
"cite_info": "Akash SR, Etha SA, et al. In vitro evaluation of antioxidant and antimicrobial properties of Cordia mocleodii leaf extract. Plant Trends. 2024; 2(2): 38-45.",
"keywords": [
"Antioxidants",
"Phytochemical",
"Alternative medicine",
"Cordia macleodii",
"Antimicrobial"
],
"DOI": "10.5455/pt.2024.04",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>Despite the rise of modern medicine, medicinal plants remain crucial in healthcare, serving as complementary therapies or potential alternatives to certain treatments [<a href=\"#r-1\">1</a>, <a href=\"#r-2\">2</a>]. This stems from the vast diversity of plant life, characterized by a rich tapestry of structural and biological variations [<a href=\"#r-3\">3</a>, <a href=\"#r-4\">4</a>]. Within plants lie unique biochemical compounds called phytochemicals, many of which possess potent inhibitory effects against pathogenic microorganisms [<a href=\"#r-5\">5</a>]. Moreover, several chronic illnesses, including diabetes, arthritis, atherosclerosis, cancer, and Alzheimer's, are linked to free radicals, highly reactive chemicals in the body [<a href=\"#r-6\">6</a>]. Evidence suggests that apoptosis and necrosis, specific cell death mechanisms, can be triggered and amplified by these free radicals and reactive nitrogen species [<a href=\"#r-7\">7</a>]. This opens doors for targeted antioxidant interventions, such as mitochondria-targeted ubiquinone, which has shown promise in alleviating liver damage from alcohol consumption [<a href=\"#r-8\">8</a>]. However, despite the potential of herbal medicine, its therapeutic capabilities remain largely unexplored, hindering its wider adoption within mainstream healthcare [<a href=\"#r-9\">9</a>, <a href=\"#r-10\">10</a>]. Therefore, a pressing need exists to systematically evaluate the efficacy of these botanical-based remedies.</p>\r\n\r\n<p><em>Cordia macleodii</em>, an endangered medicinal plant belonging to the Boraginaceae family (known locally as Dahipalas/Dahiman), is indigenous to the moist-dry deciduous forests of Central India. This plant exhibits a wide array of pharmaceutical potentials, including hepatoprotective, anti-inflammatory, antioxidant, antibacterial, antifungal, wound healing, and anti-venom activities [<a href=\"#r-11\">11</a>, <a href=\"#r-12\">12</a>]. Preliminary phytochemical analysis of plant leaves using various solvents revealed the presence of tannins, phenols, flavonoids, saponins, alkaloids, and glycosides [<a href=\"#r-13\">13</a>, <a href=\"#r-14\">14</a>]. The growing interest in natural remedies within traditional medicine systems further underscores the significance of <em>Cordia macleodii</em> and its promising therapeutic properties [<a href=\"#r-15\">15</a>].</p>\r\n\r\n<p>Screening of natural antioxidant and antibacterial properties in <em>Cordia macleodii</em> is essential for several compelling reasons. First, <em>Cordia macleodii</em> has a long history of use in traditional folk medicine. Studies have demonstrated the plant's safety and effectiveness as an alternative medicinal agent, which could legitimize and broaden its application in contemporary medicine [<a href=\"#r-16\">16</a>]. Antioxidants play a crucial role in combating oxidative stress, which is associated with various chronic illnesses, including cancer, heart disease, and neurological conditions. By identifying and evaluating the antioxidant capabilities of <em>Cordia macleodii</em>, researchers can assess its potential in managing or preventing these illnesses [<a href=\"#r-17\">17</a>].</p>\r\n\r\n<p>Moreover, the increasing prevalence of antibiotic-resistant microorganisms necessitates the discovery of new antimicrobial agents. Screening of <em>Cordia macleodii</em> for antibacterial properties may reveal compounds effective against resistant strains, contributing to the development of new antibiotics [<a href=\"#r-18\">18</a>]. The findings from antioxidant and antibacterial screenings can also enhance the creation of nutraceutical products. These food-based products can be marketed as natural health supplements, appealing to consumers interested in holistic health solutions that go beyond basic nutrition [<a href=\"#r-19\">19</a>]. Screening <em>Cordia macleodii</em> for antioxidant and antibacterial properties has great significance in therapeutic potential, combating illnesses, addressing antibiotic resistance, developing dietary supplements, and supporting conservation efforts. The aim of this study was to screen natural antioxidant and antimicrobial properties of <em>Cordia mocleodii</em> leaf extract. We evaluated the bioactive compound, and antimicrobial performance using disk diffusion assay. Initially, we have confirmed the presence of several secondary metabolites and the antibacterial potential of <em>Cordia mocleodii.</em></p>"
},
{
"section_number": 2,
"section_title": "MATERIALS AND METHODS",
"body": "<p><strong>Plant collection and extract preparation </strong></p>\r\n\r\n<p><em>Cordia macleodii </em>leaves were gathered from Savar, Dhaka, Bangladesh, and meticulously cleaned to remove extraneous plant material. After sun-drying for a week, the leaves were pulverized into a coarse powder and securely stored in airtight containers under controlled conditions. Subsequently, 180 g of the powder was soaked in 1000 ml of 90% methanol for ten days in a sealed container, with regular agitation. The resulting solution was filtered through cotton cloth and Whatman filter paper, and the filtrate was then evaporated to obtain the crude extract. This carefully collected and processed extract formed the basis for subsequent analyses, and this experiment was done three times to get the result perfectly.</p>\r\n\r\n<p><strong> </strong></p>\r\n\r\n<p><strong>Chemicals used during the experiment</strong></p>\r\n\r\n<p>This research utilized methanol, carbon tetrachloride, dichloromethane, petroleum ether, sodium carbonate, Folin-Ciocalteu reagent, quercetin, and gallic acid, all obtained from Merck Co. in Germany. Additionally, ascorbic acid was sourced from the same supplier. 1,1-Diphenyl-2-picrylhydrazyl (DPPH) was obtained from Sigma-Aldrich (United States), while standard ciprofloxacin discs were acquired from a local pharmacy. Furthermore, the streptokinase drug was supplied by Incepta Pharmaceuticals Ltd., Bangladesh.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Test microorganisms</strong></p>\r\n\r\n<p>The research utilized ten bacterial strains, comprising five Gram-positive (<em>Bacillus cereus</em>, <em>Bacillus megaterium</em>, <em>Bacillus subtilis</em>, <em>Sarcina lutea</em>, and <em>Staphylococcus aureus</em>) and five Gram-negative (<em>Escherichia coli</em>, <em>Salmonella paratyphi</em>, <em>Salmonella typhi</em>, <em>Shigella boydii</em>, and <em>Vibrio mimicus</em>) strains. These were obtained from the Institute of Nutrition and Food Science at the University of Dhaka in Bangladesh for the antimicrobial assay evaluation.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Phytochemical screening </strong></p>\r\n\r\n<p>Standard chemical assays facilitated the identification of various phytochemical groups in the samples based on their characteristic color changes. Specifically, lead acetate, alkaline reagent, ferric chloride, and ammonia were utilized to identify flavonoids and phenolics [<a href=\"#r-20\">20</a>]. Dragendroff's and Mayer's tests confirmed the presence of alkaloids, employing lead acetate, potassium hydroxide, ferric chloride, and potassium dichromate. Additionally, tannins were detected using lead acetate, potassium hydroxide, ferric chloride, and potassium dichromate. Furthermore, glycosides were identified through the Legal, Keller-Kiliani, and Borntrager tests, while saponins were detected using the froth and Salkowski tests. Finally, the Molisch test was employed to quantify the carbohydrate content.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Measurement of total flavonoids content</strong></p>\r\n\r\n<p>The <em>Cordia macleodii</em> extract was examined for its total flavonoid content (TFC) employing the aluminum chloride colorimetric method [<a href=\"#r-21\">21</a>]. A solution comprising aluminum chloride, potassium acetate, and the extract was prepared and diluted with distilled water. The absorbance of this solution was measured at a specific wavelength and compared to a blank solution lacking the extract. By employing a standard solution of quercetin, the quantity of flavonoids in the extract was determined and expressed in terms of quercetin equivalents.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Determination of total phenolic content</strong></p>\r\n\r\n<p>The quantification of total phenolic compounds (TPC) in <em>Cordia macleodii</em> extracts was measured utilizing the Folin-Ciocalteu method [<a href=\"#r-22\">22</a>] with gallic acid serving as the standard (0-100 μg/ml). In brief, 5 ml of methanol containing either 10 mg crude extract or a 2 mg/ml extract aliquot was combined with 2.5 ml of Folin-Ciocalteu reagent and 2.0 ml of 7.5% (w/v) Na2CO3 solution, then incubated at room temperature for 20 minutes. To determine the total phenolic content in the samples, absorbance at 760 nm was measured using a UV-vis spectrophotometer from Shimadzu, Japan. Researchers constructed a calibration curve based on absorbance readings of known gallic acid solutions, enabling them to express the results as milligrams of gallic acid equivalents (GAE) per gram of extract.</p>\r\n\r\n<p><strong> </strong></p>\r\n\r\n<p><strong>DPPH test for free radical scavenging activity</strong></p>\r\n\r\n<p>We utilized the Brand-Williams et al., technique to assess the plant fractions' ability to neutralize the specific free radical DPPH [<a href=\"#r-23\">23</a>] with ascorbic acid serving as the standard reference compound. Stock solutions of ascorbic acid and each fraction (1000 μg/ml) were prepared in methanol and serially diluted to obtain concentrations ranging from 0.977 to 500 μg/ml. A DPPH solution (5 mg/250 ml methanol) was prepared, and 3.0 ml aliquots were combined with 2.0 ml aliquots of each concentration of standard or fraction solution. The extent of the reaction in each mixture was evaluated by measuring its absorbance at 517 nm following a 30-minute incubation at room temperature in the absence of light, with methanol serving as a control. The percentage inhibition of DPPH radical scavenging was determined using the formula:</p>\r\n\r\n<p>% inhibition = [1 - (A sample/A blank)] × 100</p>\r\n\r\n<p>Where, A sample represents the light absorption of a solution containing the standard or fraction, and A blank represents the background absorption solely from the reagents used in the measurement. The 50% inhibitory concentration (IC50) for DPPH radical scavenging was then determined from a percentage inhibition vs. concentration curve.</p>\r\n\r\n<p><strong> </strong></p>\r\n\r\n<p><strong>Measurement of antimicrobial activity </strong></p>\r\n\r\n<p>The effectiveness of the extract against various bacteria, including both Gram-positive and Gram-negative types, was assessed using a disc diffusion method as described by Bauer et al., [<a href=\"#r-24\">24</a>]. These bacteria included <em>Bacillus cereus</em>, <em>B. megaterium</em>, <em>B. subtilis</em>, <em>Sarcina lutea</em>, <em>Staphylococcus aureus</em>, <em>Escherichia coli</em>, <em>Salmonella paratyphi</em>, <em>S. typhi</em>, <em>Shigella boydii</em>, and <em>Vibrio mimicus</em>. Bacterial strains were obtained from the Institute of Nutrition and Food Science (INFS), University of Dhaka. Sterilized 6 mm filter paper discs were impregnated with 400 μg of extract dissolved in 10% v/v DMSO. These discs, along with blank discs as negative controls and ciprofloxacin discs (5 μg/disc) as positive controls, were placed on nutrient agar plates pre-inoculated with the respective test microorganisms. Following a 24-hour incubation at 4°C to enable diffusion of the test substance, bacterial growth was assessed via a subsequent 24-hour incubation at 37°C. The antimicrobial activity of the extract was determined by measuring the diameter of inhibition zones surrounding the sample discs.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Statistical analysis</strong></p>\r\n\r\n<p>The statistical analysis of the experiments' data is presented as mean ± SEM. One-way analysis of variance (ANOVA) with Dunnett's multiple comparisons test was used to assess differences between groups. A p-value less than 0.05 was considered statistically significant and the statistical software R was used for all analyses.</p>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p><strong>Presence of phytochemicals in <em>Cordia macleodii</em> extract </strong></p>\r\n\r\n<p>Phytochemical screening unveiled the presence of several pharmacologically active constituents in the leaf extracts, including phenolics, saponins, flavonoids, tannins, glycosides, alkaloids, and carbohydrates (<a href=\"#Table-1\">Table 1</a>).</p>\r\n\r\n<p><strong> </strong></p>\r\n\r\n<p><strong>Effect of <em>Cordia macleodii</em> extract on the free radical scavenging activity</strong></p>\r\n\r\n<p>DPPH is commonly employed as a reagent to assess the antioxidant capability of substances in absorbing free radicals [<a href=\"#r-25\">25</a>]. This free radical can be stabilized by the introduction of a donor radical, such as an electron or a hydrogen radical [<a href=\"#r-26\">26</a>]. The <em>Cordia macleodii</em> extract exhibited robust antioxidant activity, demonstrated by its increasing efficacy in neutralizing DPPH radicals with rising concentration (<a href=\"#figure1\">Figure 1</a>). This effect displayed a strong positive correlation (r ~ 0.7863) with concentration, indicating a nearly perfect linear relationship. The IC50 value of <em>Cordia macleodii</em> extract was 523.21 μg/ml compared to the ascorbic acid 18.35 μg/ml of leaf extract (<a href=\"#Table-2\">Table 2</a>).</p>\r\n\r\n<div id=\"#figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"286\" src=\"/media/article_images/2024/05/30/280-1715253508-Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1.</strong> Standard carve of ascorbic acid and <em>Cordia macleodii </em>leaves extract. The figure shows the DPPH activity of <em>Cordia macleodii</em> extract with a standard antioxidant ascorbic acid.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<div id=\"Table-1\">\r\n<p><a href=\"https://plant-trends.bsmiab.org/table/280-1715253508-table1/\">Table-1</a><strong>Table 1. </strong>Presence/absence of key phytochemical constituents in <em>Cordia mocleodii</em> leaves.</p>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<div id=\"Table-2\">\r\n<p><a href=\"https://plant-trends.bsmiab.org/table/280-1715253508-table2/\">Table-2</a><strong>Table 2. </strong>Comparative analysis of DPPH inhibition by ascorbic acid and <em>Cordia macleodii</em>.</p>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Effect of <em>Cordia macleodii</em> extract on bacterial growth</strong></p>\r\n\r\n<p>The antimicrobial activity of <em>Cordia macleodii</em> leaf extract was evaluated using agar disk diffusion assay. The bacterial strains showed various potential against the pathogenic bacterial strains including gram-positive (<em>B. megaterium</em>, <em>B. subtilis</em>, <em>S. aureus</em>, <em>B. cereus</em>, <em>S. lutea</em>) and five gram-negative (<em>S. typhi</em>, <em>E. coli</em>, <em>V. mimicus</em>, <em>S. paratyphi</em>, <em>S. boydii</em>) strains (<a href=\"#figure2\">Figure 2</a>). Notably, the methanol extract exhibited potent antibacterial activity against <em>S. typhi</em>, <em>E. coli</em>, and <em>S. boydii</em> as evidenced by significantly larger inhibition zones compared to other tested microbes. Additionally, <em>B. megaterium </em>bacteria showed a higher zone of inhibition compared to other gram-positive bacteria.</p>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"293\" src=\"/media/article_images/2024/05/30/280-1715253508-Figure2.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 2. </strong>The diameter of the zone of inhibition where the <em>Cordia macleodii</em> extract showed less antibacterial activity compared to the ciprofloxacin.</figcaption>\r\n</figure>\r\n</div>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p>The vast array of bioactive compounds found in natural products has sparked significant interest due to their proven efficacy in treating various ailments. Plants, in particular, are a treasure trove of novel chemicals with diverse therapeutic applications. For example, tannins exhibit well-documented anti-inflammatory and anticancer properties [<a href=\"#r-27\">27</a>, <a href=\"#r-28\">28</a>]. Additionally, the complex interplay of secondary metabolites in medicinal plants can lead to multiple mechanisms of antimicrobial action, potentially hindering the development of resistance [<a href=\"#r-29\">29</a>]. This study investigated the antibacterial and antioxidant potential of extracts derived from <em>Cordia macleodii</em> leaves. The findings demonstrate that the extracts possess broad-spectrum antibacterial activity against various microorganisms, potentially exceeding the efficacy of conventional antibiotics. This aligns with the growing interest in natural products as a source of novel antimicrobials due to their complex composition, which can hinder resistance development [<a href=\"#r-30\">30</a>]. The observed activity might be attributed to the identified secondary metabolites, including alkaloids, terpenoids, saponins, phenols, tannins, and steroids, all of which have documented antimicrobial properties [<a href=\"#r-28\">28</a>]. Interestingly, this finding differs slightly from previous studies on <em>Cordia macleodii</em>, which reported the presence of flavonoids alongside the detected metabolites [<a href=\"#r-31\">31</a>].</p>\r\n\r\n<p>The DPPH free radical scavenging assay revealed potential antioxidant activity in the methanol extract. This aligns with the established role of phenolic compounds, such as tannins, in scavenging free radicals and mitigating oxidative stress, a key player in various diseases [<a href=\"#r-32\">32</a>, <a href=\"#r-33\">33</a>]. While previous research suggests a positive correlation between <em>Cordia macleodii</em> extract concentration and its antioxidant capacity [<a href=\"#r-34\">34</a>], this study focused on a single concentration. Future studies could explore a dose-dependent response to further elucidate the antioxidant potential. The identified phytochemicals, including glycosides, alkaloids, flavonoids, and saponins, are known contributors to various biological activities in plants [<a href=\"#r-35\">35-37</a>]. Notably, these compounds have been linked to potential thrombolytic effects, warranting further investigation into this specific activity of <em>Cordia macleodii</em> extracts. Isolating and evaluating the individual roles of these compounds would provide a deeper understanding of their contribution to the observed functionalities.</p>"
},
{
"section_number": 5,
"section_title": "CONCLUSION",
"body": "<p>This study revealed promising antibacterial, and antioxidant properties within extracts from <em>Cordia macleodii</em> leaves. Additionally, the extracts demonstrated promising antibacterial action against a variety of microorganisms, occasionally outperforming traditional medications. These results motivate further investigation to identify the specific bioactive substances responsible for these effects, and in vivo studies and exploration of the underlying mechanisms are crucial next steps. This study highlights the potential of <em>Cordia macleodii</em> extracts as a source of natural antimicrobials and antioxidants. Further research is recommended to isolate and characterize the active compounds, explore their mechanisms of action, and investigate their potential for therapeutic applications.</p>"
},
{
"section_number": 6,
"section_title": "ACKNOWLEDGEMENT",
"body": "<p>The authors would like to acknowledge the Department of Pharmacy at Bangladesh University, Dhaka, Bangladesh for providing research facilities and assistance.</p>"
},
{
"section_number": 7,
"section_title": "AUTHOR CONTRIBUTIONS",
"body": "<p>Research design and initial draft preparation: SRA, MRHS, SAE, OI; data collection and experimental assistance: SRA, MRHS, EH; data analysis: SRA, MNU, OI; manuscript review and editing: LJ. The final version of the manuscript was approved by all authors. All authors provided consent to publish this work.</p>"
},
{
"section_number": 8,
"section_title": "CONFLICTS OF INTEREST",
"body": "<p>There is no conflict of interest among the authors related to this work.</p>"
}
],
"figures": [
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/05/30/280-1715253508-Figure1.jpg",
"caption": "Figure 1. Standard carve of ascorbic acid and Cordia macleodii leaves extract. The figure shows the DPPH activity of Cordia macleodii extract with a standard antioxidant ascorbic acid.",
"featured": false
},
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/05/30/280-1715253508-Figure2.jpg",
"caption": "Figure 2. The diameter of the zone of inhibition where the Cardia macleodii extract showed less antibacterial activity compared to the ciprofloxacin.",
"featured": true
}
],
"authors": [
{
"id": 32,
"affiliation": [
{
"affiliation": "Department of Pharmacy, Bangladesh University, Dhaka-1207, Bangladesh"
}
],
"first_name": "Sajidur Rahman",
"family_name": "Akash",
"email": null,
"author_order": 1,
"ORCID": "https://orcid.org/0000-0001-8464-7570",
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 10
},
{
"id": 33,
"affiliation": [
{
"affiliation": "Department of Pharmacy, Bangladesh University, Dhaka-1207, Bangladesh"
}
],
"first_name": "Samia Alam",
"family_name": "Etha",
"email": null,
"author_order": 2,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 10
},
{
"id": 34,
"affiliation": [
{
"affiliation": "Department of Pharmacy, Bangladesh University, Dhaka-1207, Bangladesh"
}
],
"first_name": "Md. Redowan Hossain",
"family_name": "Sonet",
"email": null,
"author_order": 3,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 10
},
{
"id": 35,
"affiliation": [
{
"affiliation": "Department of Pharmacy, Bangladesh University, Dhaka-1207, Bangladesh"
}
],
"first_name": "Enamul",
"family_name": "Hoque",
"email": null,
"author_order": 4,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 10
},
{
"id": 36,
"affiliation": [
{
"affiliation": "Department of Pharmacy, Bangladesh University, Dhaka-1207, Bangladesh"
}
],
"first_name": "Md. Sujunur",
"family_name": "Rahman",
"email": null,
"author_order": 5,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 10
},
{
"id": 37,
"affiliation": [
{
"affiliation": "Department of Biochemistry and Molecular Biology, Tejgaon College, Dhaka, Bangladesh"
}
],
"first_name": "Md. Nasir",
"family_name": "Uddin",
"email": null,
"author_order": 6,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 10
},
{
"id": 38,
"affiliation": [
{
"affiliation": "Department of Pharmacy, Bangladesh University, Dhaka-1207, Bangladesh"
}
],
"first_name": "Ohidul",
"family_name": "Islam",
"email": null,
"author_order": 7,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 10
},
{
"id": 39,
"affiliation": [
{
"affiliation": "Department of Pharmacy, Bangladesh University, Dhaka-1207, Bangladesh"
}
],
"first_name": "Mst Lubna",
"family_name": "Jahan",
"email": "lubna.jahan@bu.edu.bd",
"author_order": 8,
"ORCID": "https://orcid.org/0000-0001-7412-993X",
"co_first_author": false,
"co_author": false,
"corresponding": true,
"corresponding_author_info": "Mst Lubna Jahan, Department of Pharmacy, Bangladesh University, Dhaka-1207, Bangladesh. Email: lubna.jahan@bu.edu.bd",
"article": 10
}
],
"views": 854,
"downloads": 125,
"references": [
{
"id": 269,
"serial_number": 1,
"reference": "Kebede GT. Phytochemical investigation and assessment of antibacterial activities of Calpurnia aurea root bark. Nat Prod Chem Res. 2021;9:383.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 270,
"serial_number": 2,
"reference": "Agidew MG. Phytochemical analysis of some selected traditional medicinal plants in Ethiopia. Bull Natl Res Cent. 2022;46(1):1-22.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 271,
"serial_number": 3,
"reference": "Ingle KP, Deshmukh AG, et al. Phytochemicals: Extraction methods, identification and detection of bioactive compounds from plant extracts. J Pharmacogn Phytochem. 2017;6(1):32-36.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 272,
"serial_number": 4,
"reference": "Gebreslassie HB, Eyasu A. Phytochemical screening of the leaves Calpurnia aurea (Ait.) Benth extract. Int J Clin Chem Lab Med. 2019;5:18-24.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 273,
"serial_number": 5,
"reference": "Melese A, Dobo B, et al. Antibacterial activities of Calpurnia aurea and Ocimum lamiifolium extracts against selected gram positive and gram-negative bacteria. Ethiop J Sci Technol. 2019;12(3):203-220.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 274,
"serial_number": 6,
"reference": "Akash SR, Hossain MI, et al. Devising a multi epitope vaccine toward the dengue virus using the computational method in Bangladesh. J Adv Biotechnol Exp Ther. 2023;6(1):44-57.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 275,
"serial_number": 7,
"reference": "Chatterjee S, et al. Oxidative stress induces protein and DNA radical formation in follicular dendritic cells of the germinal center and modulates its cell death patterns in late sepsis. Free Radic Biol Med. 2011;50(8):988-999.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 276,
"serial_number": 8,
"reference": "Shepard B. Antioxidant may prevent alcohol-induced liver disease. UAB News. 2011.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 277,
"serial_number": 9,
"reference": "World Health Organization. World health Organization traditional medicine strategy 2014–2023. Geneva: World Health Organization; 2013.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 278,
"serial_number": 10,
"reference": "Mulatu G. Antibacterial activities of Calpurnia aurea against selected animal pathogenic bacterial strains. Adv Pharmacol Pharm Sci. 2020;2020:1-9.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 279,
"serial_number": 11,
"reference": "Nariya PB, Shukla V, et al. Phytochemical screening and in vitro evaluation of free radical scavenging activity of Cordia macleodii bark (HOOK. F. & THOMSON). Free Radicals Antioxid. 2012;2(3):36-40.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 280,
"serial_number": 12,
"reference": "Chandrakar J, Dixit A. Cordia macleodii Hook f. Thomson-A potential Medicinal Plant. Int J Phytomedicine. 2017;9(3):394-398.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 281,
"serial_number": 13,
"reference": "Kumar V, Tiwari SS, et al. Pharmacognostic and phytochemical evaluation of Cordia macleodii. J Med Aromat Plant Sci. 2011;33(1):59-63.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 282,
"serial_number": 14,
"reference": "Upadhyay RA, Kanungo NI. Phytochemical screening and antimicrobial study of Cordia macleodii Hook F. and Thoms. on some human pathogens. Life Sci Int Res J. 2015;2347-8691.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 283,
"serial_number": 15,
"reference": "Chandrakar J, Dubey S, et al. In vitro antioxidant activity, total phenolic contents and phytochemical evaluation from crude extracts of an endangered plant Cordia macleodii Hook. f. & Thomson. J Biomed Pharm Res. 2019;8(3):111-122.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 284,
"serial_number": 16,
"reference": "Michielin EM, Salvador AA, et al. Chemical composition and antibacterial activity of Cordia verbenacea extracts obtained by different methods. Bioresour Technol. 2009;100(24):6615-6623.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 285,
"serial_number": 17,
"reference": "Valko M, Leibfritz D, et al. Free radicals and antioxidants in normal physiological functions and human disease. Int J Biochem Cell Biol. 2007;39(1):44-84.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 286,
"serial_number": 18,
"reference": "Davies J, Davies D. Origins and evolution of antibiotic resistance. Microbiol Mol Biol Rev. 2010;74(3):417-433.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 287,
"serial_number": 19,
"reference": "Bagchi D, Preuss HG. Phytopharmaceuticals in Cancer Chemoprevention. CRC Press; 2004.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 288,
"serial_number": 20,
"reference": "Ghani A. Practical phytochemistry. Dhaka: Parash Publishers; 2005.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 289,
"serial_number": 21,
"reference": "Mahmud I, Zilani MNH, et al. Bioactivities of Bruguiera gymnorrhiza and profiling of its bioactive polyphenols by HPLC-DAD. Clin Phytoscience. 2017;3(1):1-11.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 290,
"serial_number": 22,
"reference": "Škerget M, Kotnik P, et al. Phenols, proanthocyanidins, flavones and flavonols in some plant materials and their antioxidant activities. Food Chem. 2005;89(2):191-198.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 291,
"serial_number": 23,
"reference": "Brand-Williams W. Use of a free radical method to evaluate antioxidant activity. Food Sci Technol. 1999;28:1231-1237.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 292,
"serial_number": 24,
"reference": "Bauer A, Kirby W, et al. Antibiotic susceptibility testing by a standardized single disk method. Am J Clin Pathol. 1966;45(4_ts):493-496.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 293,
"serial_number": 25,
"reference": "Prasad S, Kashyap RS, et al. Effect of Fagonia arabica (Dhamasa) on in vitro thrombolysis. BMC Complement Altern Med. 2007;7(1):1-6.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 294,
"serial_number": 27,
"reference": "Oyaizu M. Studies on products of browning reaction antioxidative activities of products of browning reaction prepared from glucosamine. Jpn J Nutr Diet. 1986;44(6):307-315.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 295,
"serial_number": 28,
"reference": "Alam MM, Akash SR. In vitro pharmacological activities of methanol extract of Acmella oleracea leaves: a variety grown in Dhaka, Bangladesh. Indian J Microbiol Res. 2023;10(1):43-49.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 296,
"serial_number": 28,
"reference": "Olajide OA, Aderogba MA, et al. Effects of Anacardium occidentale stem bark extract on in vivo inflammatory models. J Ethnopharmacol. 2004;95(2-3):139-142.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 297,
"serial_number": 29,
"reference": "Ogunleye D, Ibitoye S. Studies of antimicrobial activity and chemical constituents of Ximenia americana. Trop J Pharm Res. 2003;2(2):239-241.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 298,
"serial_number": 30,
"reference": "Keita K, Darkoh C, et al. Secondary plant metabolites as potent drug candidates against antimicrobial-resistant pathogens. SN Appl Sci. 2022;4(8):209.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 299,
"serial_number": 31,
"reference": "Nigussie D, Davey G, et al. Antibacterial activity of methanol extracts of the leaves of three medicinal plants against selected bacteria isolated from wounds of lymphoedema patients. BMC Complement Med Ther. 2021;21:1-10.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 300,
"serial_number": 32,
"reference": "Usman R, Rabiu U. Antimicrobial activity of Lawsonia inermis (henna) extracts. Bayero J Pure Appl Sci. 2018;11(1):167-171.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 301,
"serial_number": 33,
"reference": "Khaliq FA, Rehman A, et al. Formulation, characterization and evaluation of in vivo wound healing potential of Lawsone ointment. Am J Adv Drug Deliv. 2018;6(01):61-68.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 302,
"serial_number": 34,
"reference": "Kachhwaha P, Gehlot HS. Changes in phytonutrients and antioxidant properties of Cordia myxa and Carissa carandas fruit during ripening. Indian J Plant Physiol. 2015;20:72-78.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 303,
"serial_number": 35,
"reference": "Sultana B, Anwar F, et al. Antioxidant activity of phenolic components present in barks of Azadirachta indica, Terminalia arjuna, Acacia nilotica, and Eugenia jambolana Lam. trees. Food Chem. 2007;104(3):1106-1114.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 304,
"serial_number": 36,
"reference": "Ali MS, Amin MR, et al. In vitro antioxidant, cytotoxic, thrombolytic activities and phytochemical evaluation of methanol extract of the A. philippense L. leaves. Asian Pac J Trop Biomed. 2013;3(6):464-469.",
"pmc": null,
"DOI": null,
"article": 10
},
{
"id": 305,
"serial_number": 37,
"reference": "Vital PG, Rivera WL. Antimicrobial activity, cytotoxicity, and phytochemical screening of Voacanga globosa (Blanco) Merr. leaf extract (Apocynaceae). Asian Pac J Trop Med. 2011;4(10):824-828.",
"pmc": null,
"DOI": null,
"article": 10
}
]
},
{
"id": 9,
"slug": "280-1715073426",
"featured": false,
"slider": false,
"issue": "Vol2 Issue2",
"type": "review_article",
"manuscript_id": "280-1715073426",
"recieved": "2024-05-26",
"revised": null,
"accepted": "2024-06-16",
"published": "2024-06-28",
"pdf_file": "https://plant-trends.bsmiab.org/media/pdf_file/2024/40/280-1715073426.pdf",
"title": "Coordination of elicitors and ascorbate-glutathione cycle: A vital nexus for mitigating post-harvest injury and losses of cultivated fruits",
"abstract": "<p>Fruit losses and waste are predominantly caused by postharvest stresses, with their management traditionally reliant on synthetic compounds. The extensive utilization of these compounds can cause harmful impacts on both human health and the environment, necessitating a reduction in their use. Recent advancements have led to the development of novel, benign strategies using natural or biologically derived compounds. These compounds, known as 'elicitors,' have the strength to instigate the plant's defense mechanism, enhancing the fruit’s resilience counter to future stress and reducing postharvest spoilage. In recent years, the impact of the plant defense system concerning elicitor compounds has been measurable through physiological parameters or the enzymatic activity of molecular markers. Antioxidant activity, a crucial parameter to determine fruit quality, shelf-life, and tolerance to biotic and abiotic stresses, significantly functions in stress response and maintaining postharvest quality. Elicitors can prompt endurance in fruits towards diverse environmental stresses by promoting the biosynthesis of antioxidant components and enhancing antioxidant capability through the ascorbate-glutathione (AsA-GSH) cycle. Recent findings have exhibited that applying chemical elicitors can enhance antioxidant activity by augmenting the content of AsA and GSH to regulate redox stability, thereby maintaining quality in harvested fruits. This review summarises recent findings demonstrating how elicitor spray can enrich antioxidant activity in harvested fruits by accelerating the synthesis of antioxidants by activating other defense mechanisms providing insight into fruit defense mechanisms induced under various elicitors and their prospects in postharvest biology.</p>",
"journal_reference": "Plant Trends. 2024; 2(2): 24-37",
"academic_editor": "Monirul Islam, PhD; University of Massachusetts Amherst Amherst, USA",
"cite_info": "Hoque MIU, Jony ME, et al. Coordination of elicitors and ascorbate-glutathione cycle: A vital nexus for mitigating post-harvest injury and losses of cultivated fruits. Plant Trends. 2024; 2(2): 24-37.",
"keywords": [
"Post-harvest injury",
"AsA-GSH cycle",
"Antioxidants",
"Chemical elicitors",
"Fruits."
],
"DOI": "10.5455/pt.2024.03",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>Fruits are abundant in beneficial compounds, playing a pivotal role in a nutritious diet for the overall well-being of human consumption. They are often harvested when green mature to minimize postharvest losses from texture softening, desiccation, and pathogen infection. However, research shows that fruits ripened naturally have better quality, especially in aroma, than those ripened postharvest [<a href=\"#r-1\">1, 2</a>]. Fruits are highly perishable, and various factors can impact their original attributes. Global fruit post-harvest losses are estimated between 25 and 50% of total production. Even under ideal storage conditions, they are susceptible to decomposing during the post-harvest period [<a href=\"#r-3\">3</a>]. A recent FAO report highlighted that fruits have the second highest value of losses and waste (~22%) in the food supply chain, after roots, tubers, and oil-bearing crops, due to their perishable nature [<a href=\"#r-4\">4</a>]. Metabolism of Postharvest fruits is influenced by endogenous (intrinsic physiology) and exogenous factors (abiotic and biotic). Ripening and senescence can vary based on genotype through interaction with these factors [<a href=\"#r-5\">5, 6</a>]. In response, inorganic chemical substances have been traditionally used to manage these stresses during the postharvest period. Their frequent use has led to the development of microbial resistance and pesticide residues in fruits, which poses health risks raising environmental concerns.<br />\r\nSeveral government policies along with the Food and Agriculture Organization (FAO) recommended limiting the use of these inorganic harmful pesticides in crops. Research is ongoing to identify cost-effective, safe, environmentally friendly alternatives for controlling fruit diseases. Elicitors or resistance signaling inducers have been a focus of scientific research for over two decades as potential replacements for pesticides [<a href=\"#r-7\">7, 8</a>].<br />\r\nFruits can experience oxidative stress as they ripen and undergo senescence after harvest, which results in higher production of ROS, which are natural by-products of metabolism, growth and development. They are distributed across various organelles within the fruit cells. Plants have developed nonenzymatic and enzymatic mechanisms to regulate the balance between ROS generation and scavenging under normal physiological conditions. However, specific abiotic stressors can produce excessive ROS during oxidative stress, resulting in collateral destruction of cell structures, membrane lipid peroxidation, and impairment of proteins and DNA, which results in abnormal fruit ripening. Early responses in postharvest fruit are often due to disruptions in the system that produces and scavenges ROS [<a href=\"#r-9\">9</a>]. Antioxidants like ascorbate (AsA) and glutathione (GSH) play a key role in combating ROS, along with superoxide dismutase (SOD), peroxidase (POD), catalase (CAT), and ascorbate peroxidase (APX). The AsA-GSH cycle is crucial for ROS removal during postharvest storage. Fruits with sufficient levels of GSH, AsA, APX, GR, MDHAR, and DHAR involved in the AsA-GSH pathway exhibit improved tolerance to oxidative stress. Numerous investigations have focused on the importance of the AsA-GSH cycle in balancing the postharvest standard of fruits and holding up their senescence with elicitors (physical/chemical) application like plant hormones, nanoparticles, compounds gas, ray, etc., which also can enhance the ROS scavenging power, reducing ROS amount in host fruits to mitigate fruit damage and coordinate the induction of other defense response regulatory pathways [<a href=\"#r-1\">1</a>, <a href=\"#r-7\">7</a>, <a href=\"#r-8\">8</a>, <a href=\"#r-10\">10-17</a>]. This review highlights recent advancements in using different elicitors on fruits after harvest to enhance the stress-reducing AsA-GSH cycle. Additionally, it explores how these elicitors activate diversified mechanisms with relevant molecular pathways in fruits, providing insights into the broader context of plant defense networks.</p>"
},
{
"section_number": 2,
"section_title": "POST-HARVEST STRESS IN FRUITS: ROS, A KEY FACTOR",
"body": "<p>Regular consumption of fresh fruits provides essential nutrients and antioxidants that help reduce the risks of lifestyle-related diseases. However, after harvesting fruits and vegetables, their storage duration, and shelf-life are important for the agriculture industry. Managing postharvest fruits involves balancing ROS and antioxidants, which can be challenging due to the plant’s response to stress signals. Excessive ROS levels can damage cells and proteins, leading to lipid peroxidation. Maintaining a sustainable balance between ROS and antioxidants is important to prevent stress in postharvest produce. Signalling chemicals alternatively known as elicitors can help increase antioxidant production and protect against postharvest handling and distribution. This can be achieved by upregulating the Ascorbate-Glutathione (AsA-GSH) cycle [<a href=\"#r-7\">7</a>, <a href=\"#r-8\">8</a>, <a href=\"#r-18\">18</a>].</p>"
},
{
"section_number": 3,
"section_title": "ROLE OF AsA-GSH CYCLE IN MITIGATING ROS-MEDIATED STRESS",
"body": "<p>ROS is produced during aerobic reactions when oxygen is partially reduced or activated. ROS are a group of reactive components that include “superoxide (O<sub>2</sub><sup>−</sup>), hydroxyl (OH•), and peroxyl (ROO•), as well as nonradical forms like hydrogen peroxide (H<sub>2</sub>O<sub>2</sub>), singlet oxygen (<sup>1</sup>O<sub>2</sub>), and ozone (O<sub>3</sub>). ROS play crucial dual functions in plants: at moderate levels, they work as signaling molecules to induce responses to abiotic stress for adaptation, while excessive amounts can cause oxidative bursts. At the highest point, this oxidative damage can lead to lipid peroxidation of membranes and protein and nucleic acid degradation, potentially culminating in the oxidative destruction of the cell. ROS are produced in several cellular locations, including the chloroplast, mitochondrion, cell membranes, and apoplast. Upon initial stress perception, plants synthesize specific hormones (salicylic acid—SA, jasmonic acid—JA, ethylene—ET, and abscisic acid—ABA) tailored to the required physiological response. These hormones propagate through the plant system, initiating signaling pathways that modulate gene expression changes. Consequently, antioxidant enzymes are synthesized to scavenge the elevated ROS levels induced by oxidative stress, thus supporting redox potential and plant survival under stress conditions. The cross-talk among these components improves signaling relations and stabilizes the plant defense system. [<a href=\"#r-19\">19</a>]. Plant’s antioxidants in organelles neutralize the ROS. Excessive amounts of ROS production can cause oxidative damage [<a href=\"#r-20\">20</a>, <a href=\"#r-21\">21</a>].</p>"
},
{
"section_number": 4,
"section_title": "BIOSYNTHESIS OF ASCORBATE AND GLUTATHIONE",
"body": "<p>AsA biosynthesis, one of the most ancient biological pathways, is crucial for various primitive life processes on Earth. In plants, AsA is mainly produced via the L-galactose pathway, also referred to as the Smirnoff–Wheeler pathway. This pathway begins with converting D-glucose-6-phosphate to D-fructose-6-phosphate, followed by a sequence of enzymatic reactions that culminate in the synthesis of AsA. The pathway's key enzymes include phosphoglucose isomerase (PGI), phosphomannose isomerase (PMI), and GDP-mannose pyrophosphorylase (GMP), among others. Notably, the enzyme L-galactono-1,4-lactone dehydrogenase (GLDH), crucial in the final step, is located in the mitochondrial inner membrane, indicating mitochondrial involvement in AsA synthesis. While the L-galactose pathway is predominant, alternative pathways such as the L-gulose and D-galacturonic acid pathways also contribute to AsA production, underscoring the complexity and versatility of AsA biosynthesis linked to cell wall formation [<a href=\"#r-20\">20</a>, <a href=\"#r-21\">21</a>].<br />\r\nGlutathione (GSH), a ubiquitous tripeptide composed of glutamine, cysteine, and glycine, is a potent antioxidant and a vital metabolite with diverse roles in plants, ranging from antioxidant defense to cell cycle regulation. GSH synthesis occurs in two ATP-dependent enzymatic steps, starting with γ-glutamylcysteine formation, followed by its conversion to GSH, facilitated by GSH synthetase. GSH is crucial for cellular antioxidant capacity, particularly under abiotic stress, and AsA regeneration in the AsA-GSH cycle. This cycle is integral for maintaining cellular redox homeostasis and enabling plants to withstand various stress conditions [<a href=\"#r-20\">20-22</a>].</p>"
},
{
"section_number": 5,
"section_title": "ENZYMES OF AsA-GSH INVOLVED IN REDOX SIGNALISING",
"body": "<p>AsA/DHA and GSH/GSSG play vital roles as redox managers in maintaining the redox balance within plant cells. Ascorbate is key in scavenging ROS and cellular processes such as “electron donation to photosystem II (PSII), violaxanthin de-epoxidation, and α-tocopherol reductive quenching”. The AsA pool directly reduced MDHA by ferredoxin at photosystem I (PSI), Monodehydroascorbate reductase (MDHAR), and dehydroascorbate (DHA) reduction via GSH-dependent dehydroascorbate reductase (DHAR) activity. The redox potential of GSH is influenced by its concentration and GSH/GSSG ratio. Both AsA and GSH are essential components of the cellular redox homeostasis network. In this cycle, AsA donates electrons to ascorbate peroxidase (APX), which detoxifies H2O2 by its high affinity for H2O2, allows it to scavenge ROS effectively even at low concentrations, producing DHA, which is then recycled to maintain a high AsA/DHA ratio. If DHA is not reduced, it may be irreversibly hydrolyzed, diminishing the AsA redox pool’s efficacy. Ascorbate oxidase (AO) creates a redox gradient across the plasma membrane and facilitates apoplastic and cytoplasmic signalling. Conversely, the GSH/GSSG couple balances the intracellular redox potential and signalling. Thus, understanding the distribution of GSH across different cellular organelles is crucial for effective signalling, cellular metabolism, and fruit defense.<br />\r\nSOD catalyses the dismutation of the superoxide anion (•O<sub>2</sub><sup>−</sup>) to form H<sub>2</sub>O<sub>2</sub> and O<sub>2</sub> to counteract oxidative stress [<a href=\"#r-31\">31</a>]. SOD works in close synchrony with additional enzymes, for instance, catalase (CAT) and plant peroxidases (POX), to prevent the formation of more harmful ROS through the Haber-Weiss reaction. At low concentrations of H<sub>2</sub>O<sub>2</sub>, this enzyme stays inactive. However, when H<sub>2</sub>O<sub>2 </sub>concentrations increase, a second molecule of H<sub>2</sub>O<sub>2</sub> functions as a reductant for the enzyme producing water and oxygen as outputs. Compared to ascorbate APX, CAT has a higher Km for H<sub>2</sub>O<sub>2</sub>, making it more active at higher concentrations. APX is located in multiple subcellular compartments, allowing finely tuned scavenging activity. POXs, or class III peroxidases, function in the extracellular space to scavenge H<sub>2</sub>O<sub>2</sub>. Regarding antioxidant mechanisms, the interaction of ascorbate-glutathione molecules maintains redox balance through the Foyer-Halliwell-Asada also known as AsA-GSH pathway. Flavonoids, phenolic compounds, and POX are backup defense against excessive H<sub>2</sub>O<sub>2</sub> [<a href=\"#r-19\">19</a>, <a href=\"#r-20\">20</a>].<br />\r\nGPX catalyzes the conversion of H<sub>2</sub>O<sub>2</sub> to water and lipid alcohols, respectively. It has been considered as a biomarker for intracellular oxidative stress. Glutathione reductase (GR), a flavoprotein oxidoreductase NAD(P)H-dependent enzyme, is crucial in the AsA pathway, reducing GSSG to reduced GSH. Ascorbate peroxidases (APX), a class I heme peroxidase familiar as ascorbate-dependent peroxidase, scavenges H<sub>2</sub>O<sub>2</sub> and senses redox alterations within plant cells, playing an essential role in the “Foyer-Halliwell-Asada pathway”. It uses ascorbate to convert H<sub>2</sub>O<sub>2 </sub>to water, producing mono dehydroascorbate reductase (MDHA) as an intermediate. MDHA, which is unstable and quickly converted to DHA, is reduced and recapitulates ascorbate by MDHAR using NADH or NADPH. This enzyme's activity, reported in diverse cellular compartments, namely chloroplasts and mitochondria, is vital for maintaining a reduced ascorbate pool. DHAR breaks DHA to ascorbic acid, utilizing reduced GSH, which is critical for maintaining sufficient ascorbate levels. The continuous recycling of small soluble antioxidants like ascorbate and GSH is crucial for maintaining cellular redox balance. The antioxidant defense system's protective role extends to safeguarding unsaturated membrane lipids, nucleic acids, enzymes, and other cellular structures, making it a topic of significant interest in plant growth and development research [<a href=\"#r-19\">19</a>, <a href=\"#r-20\">20</a>].<br />\r\nFrom a recent study, GSH (5 mM) significantly mitigated weight loss in mangoes by acting as an antioxidant. When applied as an edible coating on the fruit peel, it maintains low total soluble solids (TSS), inhibiting the rapid increase in sugar content during ripening towards lower lipid peroxidation indicated by lower malondialdehyde (MDA) levels maintaining a high GSH level and favourable GSH/GSSG ratio help to regulate redox balance to oxidative stress homeostasis during ripening, and post-harvesting and storage of fruits. The impact of exogenous GSH on chilling injury (CI) in postharvest bell pepper fruits stored at low temperatures was mitigated by its role in stabilizing membranes and maintaining organelle integrity through the AsA-GSH cycle. Cold stress is mitigated by enhancing the expression of Asa-GSH cycle-related genes <em>CaGR2, CaAPX1, CaGR2, CaMDHAR1 </em>and <em>CaDHAR1</em>.</p>"
},
{
"section_number": 6,
"section_title": "ELICITORS AND AsA-GSH CYCLE FOR MITIGATING POSTHARVEST INJURY AND DAMAGES",
"body": "<p>Oxidative burst (ROS) is a common response to stress in fruits, leading to the rapid production of ROS. Induced resistance is a new approach to managing postharvest stress by activating fruit immune responses with external elicitors. This process involves signal transduction to enhance stress resistance genes and biosynthesis of secondary metabolites. Elicitors can influence the antioxidant defense system by upregulating the AsA-GSH cycle c0-ordinated other defense pathways (<a href=\"#figure1\">Figure 1</a>) (<a href=\"#Table-1\">Table 1</a>) [<a href=\"#r-23\">23</a>, <a href=\"#r-24\">24</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Chitosan</strong><br />\r\nChitosan is a natural biopolymer derived from crustaceans, insect cuticles, and fungal cell walls consisting of N-acetyl-D-glucosamine and D-glucosamine connected by β-1,4-glycosidic bonds well-known for its biocompatibility, biodegradability, antibacterial properties. Recent studies have highlighted chitosan, an elicitor of plant defense, enhancing plant resistance to multiple stresses by activating defense-related responses accumulating the antioxidants through synthesis (Phenolics, flavonoids), and upregulating defense enzymes like polyphenol oxidase (PPO), phenylalanine ammonia-lyase (PAL), nonenzymatic components, strengthening plant cell walls, providing a mechanical barrier to biotic stresses like pathogen attacks, boosting the effectiveness of defense enzymes like β-1,3-glucanase, chitinase, and chitosanase named pathogenesis-related (PR) proteins, enhance plant resistance against different pathogens by breaking down fungal wall polysaccharides and attacking invading pathogens and induces the signaling pathways of phytohormones, directly activating plant defenses against stress [<a href=\"#r-14\">14</a>, <a href=\"#r-25\">25</a>].<br />\r\nAs a sub-tropical fruit with significant business value in the world market, grapefruit (Citrus paradisi Macf.) is primarily consumed fresh. Carboxymethylation, a modification that improves the solubility of chitosan to produce water-soluble carboxymethyl chitosan (CMCS), is promising for its application in protecting fruit injury and damage. It can enhance resistance to postharvest diseases like blue and green mold in citrus fruits caused by fungi such as ‘P. expansum and P. digitatum’ by activating specific transcription factors (TFs), particularly from the AP2/ERF, WRKY, DREB, and MYB families activating hormonal signaling pathways can operate the ROS scavenging system. Increased expression of the genes involved in ABA metabolism boosted the activation of antioxidants and delaying senescence [<a href=\"#r-10\">10</a>]. Additionally, the immediate post-harvest induction of CMCS altered enzymes related to the antioxidant mechanism and AsA-GSH cycle in the case of grapefruit [<a href=\"#r-14\">14</a>, <a href=\"#r-25\">25</a>]. <br />\r\nThe cell wall is a strong barrier against pathogens and a key defense mechanism in post-harvest fruit. The cell wall’s polysaccharides, such as pectin, cellulose, and hemicellulose, create a network that gives mechanical strength to fruit tissue can be degraded, and loss of firmness during storage can be suppressed Using CMCS with significantly improved levels of GSH and AsA, However, CMCS served as an elicitor of postharvest enhancement of cell wall protection and antioxidant function needs more investigation and exploration [<a href=\"#r-14\">14</a>, <a href=\"#r-25\">25</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Carboxymethyl cellulose</strong><br />\r\nEdible coatings as polysaccharides are applied to products to preserve quality by controlling physiological, biochemical, and oxidation processes, protecting agricultural items from physio-chemical and biological issues incorporated with active ingredients, flavourings, and colorings to enhance fruit’s nutritional and sensory attributes. Carboxymethyl cellulose is a high molecular weight compound made up of glucopyranosyl units. It enhances the strength and structure of edible coatings, making them more stable and durable, and widely used in the food industry. These coatings also delay the ripening process in climacteric fruits as protective barriers against gas exchange with the environment, adjusting oxygen and water transfer rates [<a href=\"#r-20\">20</a>] [<a href=\"#r-13\">13</a>, <a href=\"#r-26\">26</a>].<br />\r\nAonla fruit improvements in preservation, when coated with a 1% CMC solution effectively, reduced the levels of H<sub>2</sub>O<sub>2</sub>, MDA, and superoxide anions in the treated fruits exhibited enhanced activities of SOD, APX, POD, and CAT, alongside the attributes related to the ASA-GSH cycle. Moreover, the coated fruits showed the highest amount of total phenolic and flavonoid contents indicated with increased phenylalanine ammonia-lyase (PAL), reduced polyphenol oxidase (PPO) showed anti-browning activities. Developing a CMC-based protective layer effectively delayed chlorophyll degradation by suppressing the activities of degrading enzymes named chlorophyllase-phytyl (Chl-Phy) and chlorophyll peroxidase (Chl-POD) enzymes [<a href=\"#r-13\">13</a>, <a href=\"#r-26\">26</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>GABA (γ-aminobutyric acid)</strong><br />\r\n(GABA) γ-aminobutyric acid is notable for its extensive literature connecting it to crucial jobs in plant stress physiology. GABA links carbon and nitrogen metabolic fluxes through the GABA shunt and influences the stress response pathways indirectly influencing ROS homeostasis by interacting with polyamines (PAs). Under stress conditions leading to an elevated NADH/NAD+ ratio, GABA can be catabolized to γ-hydroxybutyrate (GHB) may mitigate excessive ROS accumulation by supplying NADPH via the action of γ-hydroxybutyrate dehydrogenase (GHBDH), supporting detoxifying enzymes, including those in the ascorbate/glutathione cycle. Proline metabolism, closely linked with GABA’s, is also activated under stress and can be converts to glutamate by ProDH and P5CR, which can be converted to GABA is further supported by enzymatic and nonenzymatic reactions, emphasizing the intricate metabolic networking essential for plant stress adaptation (<a href=\"#Table-1\">Table 1</a>) [<a href=\"#r-24\">24,25,26</a>].<br />\r\nThe function of MIOX, a key enzyme in the myo-inositol pathway, in AsA biosynthesis remains to be determined. Various responses of PpMIOX4 transcripts to GABA treatment suggest the need for further investigation into how PpMIOX4 influences AsA biosynthesis in peach fruits [<a href=\"#r-27\">27</a>]. GABA treatment also significantly improved the expression of all three GSH biosynthetic genes -PpGR2, PpDHAR2, PpMDHAR, PpGPX2, and PpAPX of the AsA-GSH cycle in peaches, promoting GSH accumulation. The ethylene-responsive factor (ERF) subfamily, which constitutes the most significant group within the plant AP2/ERF superfamily, has been extensively studied for its roles in conferring resistance to various abiotic stresses such as drought, salinity, and chilling [<a href=\"#r-28\">28</a>] by controlling several metabolic pathways in Arabidopsis, tomato, and citrus. The GABA induced the expression of PpERF in fruits, suggesting a potential role of ERFs in activating AsA and GSH [<a href=\"#r-29\">29</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Hydrogen sulfide </strong><br />\r\nIn plant cells, hydrogen sulfide (H<sub>2</sub>S) is found in various subcellular compartments associated with enzymes involved in sulfur metabolism. The chloroplast helps in the sulfate assimilation by hosting sulfite reductase, which catalyses sulfite reduction to sulfide. Alternative pathways for sulfide generation exist in the cytosol and mitochondria. In the cytosol, H<sub>2</sub>S can be unfolded from cysteine through the action of enzymes like ‘D-cysteine desulfhydrase (DCD) and L-cysteine desulfhydrase (LCD)’, resulting in the production of ammonia and pyruvate. Meanwhile, in the mitochondria, β-cyanoalanine synthase facilitates H<sub>2</sub>S production by converting cyanide to β-cyanoalanine, utilizing cysteine. Accumulating evidence highlights H<sub>2</sub>S as a significant signaling molecule in plant-environment interactions. H<sub>2</sub>S interacts with the oxidative stress response network involving ROS at different levels, including the modulation of ROS-processing systems through changes in gene expression and protein modifications. The crosstalk between H<sub>2</sub>S and ROS also involves factors like nitric oxide, influencing crucial cellular processes. Treatment with H<sub>2</sub>S has been shown to increase the activities of several enzymes including APX, DHAR, GR, SOD, and CAT in the AsA-GSH pathway [<a href=\"#r-12\">12</a>,<a href=\"#r-27\">27,28</a>]. <br />\r\nHydrogen sulfide-applied banana fruits exhibited augmented proline (An essential osmolyte) levels and related enzyme activities [<a href=\"#r-28\">28</a>]. Hydrogen sulfide treatment increased P5CS and OAT activities and decreased PDH a rate-limiting enzyme of proline degradation activity, leading to elevated proline levels in the fruits during chilling stress. Similarly, hydrogen sulfide fumigation led to higher endogenous GABA levels and showed increased total phenolic content, attributed to decreased activity of PPO, increased PAL activity evidenced reduced browning. Furthermore, treated banana plants demonstrated elevated activities of key antioxidant enzymes. It synergistically regulates plant energy metabolism through enzymes such as H+-ATPase, Ca2+-ATPase, and cytochrome C oxidase [<a href=\"#r-30\">30</a>], resulting in delayed cellular energy loss and significantly improved chilling tolerance of banana fruits. The breakdown of chlorophyll involves specific chlorophyll catabolism enzymes. In contrast, hydrogen sulfide fumigation markedly inhibited these enzymes, thereby preserving the greenness of banana peels under postharvest chilling stress [<a href=\"#r-31\">31</a>]. Phenolic compounds are known for their ROS scavenging potential, reducing oxidative stress and attenuating membrane lipid peroxidation, thus enhancing Chilling Injury (CI) tolerance. Hydrogen sulfidic fumigation of banana fruits substantially reduced POD and PPO activities with increased PAL activity, leading to higher phenolic content, suggesting effectively mitigates CI in bananas stored under cold conditions (<a href=\"#figure1\">Figure 1</a> and <a href=\"#Table-1\">Table 1</a>) [<a href=\"#r-12\">12</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Ozone</strong><br />\r\nOzone (O<sub>3</sub>), a triatomic form of oxygen, forms when molecular oxygen (O<sub>2</sub>) reacts with an oxygen-free radical. It has a characteristic pungent smell and appears as a blue gas when generated from dry air, although it is colorless when derived from high-purity oxygen. In typical concentrations used for food processing, its color is not detectable. Ozone has a relatively high oxidation-reduction potential and is nearly 13 times more soluble in water than oxygen, making it a potent oxidizing agent that rapidly decomposes back to atmospheric oxygen, leaving no residues. Its antimicrobial activity results from direct interactions (ozone molecules) and indirect effects (free radicals). In addition to its antimicrobial properties, ozone can enhance plant stress tolerance by stimulating cellular ROS scavenging systems, thus boosting the synthesis of antioxidant enzymes. Ozone treatment influences different reactions by enhancing antioxidant enzyme activities, including POD, superoxide dismutase (SOD), and PAL while inhibiting PPO activity. Furthermore, the efficacy of O<sub>3</sub> in maintaining the quality of minimally processed products is evident through its influence on total soluble solid (TSS) content and titratable acidity (TA). Overall, Ozone acts as a signal to induce rapid defensive responses in plants, activating multiple pathways that enhance their capability to reduce free radicals, thereby prolonging the freshness of postharvest vegetables [<a href=\"#r-32\">32-34</a>].<br />\r\nOzone treatment improves postharvest storage of cantaloupe by enhancing antioxidant capacity, particularly in the ASA-GSH cycle by reducing H<sub>2</sub>O<sub>2</sub> and MDA accumulation. Additionally, mechanical processing can accelerate senescence and decay in red pitaya. Overall, low-concentration, long-term ozone treatment can delay aging and preserve the quality of fruits [<a href=\"#r-35\">35</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Methyl jasmonate</strong><br />\r\nJasmonates (JAs) are cyclopentanone components or plant hormones derived from α-linolenic acid. JA, including its derivatives like methyl jasmonate (MJ), forms a set of oxygenated fatty acids and is a critical stimulator in their biosynthesis. JA is synthesized from fatty acids in three steps, starting in the chloroplast membrane. Key enzymes related in this pathway include lipoxygenase (LOX), allene oxide synthase (AOS), allene oxide cyclase (AOC), hydroperoxide lyase (HPL), alcohol acyltransferase (AAT), and alcohol dehydrogenase (ADH), which together facilitate the synthesis of jasmonic acid and its volatile organic compounds (VOCs) in plant cells [<a href=\"#r-33\">33</a>]. Jasmonates are essential for mediating plant's biotic and abiotic stresses. This interaction triggers a series of defense responses, including the production of ROS and reactive nitrogen species (RNS) and the induction of enzymes that protect against oxidative stress also triggers the synthesis and build-up of signaling molecules like JA, SA, NO, and ET within the cell, thereby influencing the expression of genes related to secondary metabolite production. Nevertheless, MeJA is the most widely used elicitor perceived by specific receptors involved in signal transduction [<a href=\"#r-33\">33,34</a>]. Variations in the expression of genes encoding key enzymes directly influence the accumulation of corresponding secondary metabolites, with PAL regulation, MEP (methylerythritol phosphate), and MVA (mevalonate) pathways seeing increased expression with MeJA treatment [<a href=\"#r-36\">36</a>, <a href=\"#r-37\">37</a>].<br />\r\nGC-MS analysis indicated that fruits at the complete red stage contained more types and higher concentrations of VOCs compared to those at the early ripening stage. Notably, many VOCs are derived from the LOX pathway. Moreover, the content of VOCs increased with the rise in ethylene production. Correlation analysis showed a strong positive relationship between VOC content and ethylene production, suggesting that ethylene is a significant mediator in VOC biosynthesis. The application of 1-methylcyclopropene (1-MCP) appeared to decrease the accumulation of VOCs by inhibiting ethylene production, whereas MeJA treatment enhanced VOC accumulation by stimulating ethylene production. Additionally, it was found that MeJA treatment promoted VOC accumulation and regulated the antioxidant system, including the ascorbate-glutathione (AsA-GSH) cycle [<a href=\"#r-1\">1</a>].<br />\r\nResearchers are increasingly focusing on the ICE-CBF-COR signaling pathway, recognized as a critical component of the cold acclimation process. Inducer of CBF Expression (ICE), a MYC-type basic helix-loop-helix (bHLH) family transcription factor, plays a central role in the induction of C-repeat binding factors (CBFs) during cold exposure. This pathway is essential for activating the expression of downstream genes encoding osmoregulatory substances. Although the ICE-CBF-COR pathway is pivotal in plant freezing tolerance, current research also examines the influence of hormones on cold stress responses. Essential hormones such as ABA, GA, BRs, JA, auxin, CK, melatonin, and polyamines impact CBF expression [<a href=\"#r-38\">38</a>]. MeJA-treated fruits exhibited significantly increased expression levels of the C-repeat-binding factor 1 (SlCBF1), a cold-regulated gene (SlCOR413), and inducers of SlCBF expression (SlICE1 and SlICEa), all components of the SlICE-SlCBF-SlCOR (ICC) signaling pathway.<br />\r\nAdditionally, correlation analysis suggested that genes involved in the AsA-GSH cycle and the ICC signaling pathway were closely associated with the transcription of SlMYC2, a jasmonic acid signaling transcription factor. These findings underscore the role of MeJA-mediated regulation of the AsA-GSH cycle and the ICC signaling pathway during cold tolerance in tomato fruits (<a href=\"#figure1\">Figure 1</a>) (<a href=\"#Table-1\">Table 1</a>) [<a href=\"#r-10\">10</a>, <a href=\"#r-11\">11</a>].</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"355\" src=\"/media/article_images/2024/01/29/280-1715073426-Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1</strong>. A proposed working model of elicitors and AsA-GSH coordination in mitigating post-harvest fruit injury and damages. A series of elicitors induce several physiological and metabolic alterations including the ascorbate-glutathione pathway. The alteration of these processes improves post-harvest fruit defense which mitigates fruit injury and damages. </figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<div id=\"Table-1\">\r\n<p><a href=\"https://plant-trends.bsmiab.org/table/280-1715073426-table1/\">Table-1</a><strong>Table 1</strong>. Role of elicited ASA-glutathione pathway in postharvest stress mitigation in fruit.</p>\r\n</div>"
},
{
"section_number": 7,
"section_title": "CONCLUSION AND FUTURE PROSPECTS",
"body": "<p>Systemic acquired resistance (SAR) is a key trait for protecting plants or fruits from pathogen attacks. Several secondary metabolites and amino acids including carotenoid biosynthesis, accumulation of flavonoids, carotenoids, and ascorbic acid in response to elicitors are crucial. Accumulation of these metabolites along with boost-up antioxidant activity through the AsA –GSH cycle in harvested fruit, scavenges ROS maintains cellular integrity, and declines lipid peroxidation. Furthermore, the alteration of a series of metabolic changes stabilizes membrane phospholipid metabolism and maintains cell membrane potential and fluidity, which helps to preserve postharvest fruit quality. Importantly, the elicitor responses delay fruit ripening by suppressing ethylene production and respiration rates that lead to maintaining sugar levels, organic acids, and firmness during storage. Postharvest elicitor responsiveness provides a simple and cost-effective strategy with plant defense for managing harvested fruit's abiotic and biotic stresses. Additionally, the efficacy of elicitors may vary based on fruit species or cultivars, elicitor type, concentration, and timing. Agricultural practices such as fertilization, irrigation, and pruning also influence the effectiveness of elicitors. Furthermore, the application technique—including spray density and intensity—and the equipment used are critical factors. Postharvest ecological conditions, including storage temperature, relative humidity, and gas composition, also affect elicitor performance, and optimal combinations of these factors must be determined. Despite extensive research into the effects of postharvest elicitor spray on fruit properties and a preliminary understanding of the physiological and cytological mechanisms, deeper mechanisms remain elusive. Advanced multi-omic approaches and gene-editing approaches are necessary for improving fruit shelf-life, and storage duration and mitigating stress-induced damages. Moreover, elicitors-induced accumulation of various natural plant metabolites and amino acids offer safer alternatives to using chemical fungicides and lower risk of resistance development. Future research must disclose the strengthening of the linkage between pre- and postharvest periods and broaden the scope of postharvest studies by application of different combined elicitors to gain rapid effects like synthetic compounds. Fruits and other edible organs provide numerous fruitful nutrients that enrich our lives and society. Unfortunately, around 33% of harvested fruits and other edible products are never consumed due to short-shelf-life, and not well-equipped storage system that causes post-harvest waste and economic loss. However, several breeding strategies can retain desirable features by declining post-harvest damages and increasing plant Defense and fruit shelf-life. Considering the demand for valuable fruits, vegetables and other edible crops new gene editing tool would be promising and effective for fruits and other edible parts improvement by improving desirable traits. This editing approach may improve traits related to fruit quality, shelf-life, yield, and post-harvest losses of fruits. Furthermore, CRISPR–Cas9 method would be a promising tool for alternative solutions of improving traits by reducing post-harvest waste and economic loss of fruits. </p>"
},
{
"section_number": 8,
"section_title": "ACKNOWLEDGEMENTS",
"body": "<p>None.<br />\r\n </p>"
},
{
"section_number": 9,
"section_title": "AUTHOR CONTRIBUTIONS",
"body": "<p>MI and SMA: research concept initiation and first draft writing. MEJ: data collection and curation. AKD: figure drawing. SS, MMR, and IHS: writing, review, and editing. SMA: coordinated the whole work.</p>"
},
{
"section_number": 10,
"section_title": "CONFLICTS OF INTEREST",
"body": "<p>There is no conflict of interest among the authors related to this work.</p>"
}
],
"figures": [
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/01/29/280-1715073426-Figure1.jpg",
"caption": "Figure 1. A proposed working model of elicitors and AsA-GSH coordination in mitigating post-harvest fruit injury and damages. A series of elicitors induce several physiological and metabolic alterations including the ascorbate-glutathione pathway. The alteration of these processes improves post-harvest fruit defense which mitigates fruit injury and damages.",
"featured": true
}
],
"authors": [
{
"id": 25,
"affiliation": [
{
"affiliation": "Department of Applied Biosciences, Kyungpook National University, Daegu 41566, Republic of Korea"
}
],
"first_name": "Md. Injamum-Ul-",
"family_name": "Hoque",
"email": null,
"author_order": 1,
"ORCID": null,
"co_first_author": true,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 9
},
{
"id": 26,
"affiliation": [
{
"affiliation": "Department of Food Engineering, Bangabandhu Sheikh Mujibur Rahman Science and Technology University, Gopalganj-8100, Bangladesh"
}
],
"first_name": "Md. Entaduzzaman",
"family_name": "Jony",
"email": null,
"author_order": 2,
"ORCID": null,
"co_first_author": true,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 9
},
{
"id": 27,
"affiliation": [
{
"affiliation": "Department of Applied Biosciences, Kyungpook National University, Daegu 41566, Republic of Korea"
}
],
"first_name": "Ashim Kumar",
"family_name": "Das",
"email": null,
"author_order": 3,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 9
},
{
"id": 28,
"affiliation": [
{
"affiliation": "Department of Applied Biosciences, Kyungpook National University, Daegu 41566, Republic of Korea"
}
],
"first_name": "Shifa",
"family_name": "Shaffique",
"email": null,
"author_order": 4,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 9
},
{
"id": 29,
"affiliation": [
{
"affiliation": "Institute of Genomics for Crop Abiotic Stress Tolerance, Department of Plant and Soil Science, Texas Tech University, Lubbock, TX, 79409, USA"
}
],
"first_name": "Md. Mezanur",
"family_name": "Rahman",
"email": null,
"author_order": 5,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 9
},
{
"id": 30,
"affiliation": [
{
"affiliation": "Department of Biotechnology and Genetic Engineering, Bangabandhu Sheikh Mujibur Rahman Science and Technology University, Gopalganj-8100, Bangladesh"
}
],
"first_name": "Imdadul Haque",
"family_name": "Sharif",
"email": null,
"author_order": 6,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 9
},
{
"id": 31,
"affiliation": [
{
"affiliation": "Department of Plant Medicals, Andong National University, Andong, 36729, Republic of Korea"
}
],
"first_name": "S.M.",
"family_name": "Ahsan",
"email": "smvahsan@gmail.com",
"author_order": 7,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": true,
"corresponding_author_info": "S.M. Ahsan, Department of Plant Medicals, Andong National University, Andong, 36729, Republic of Korea.\r\nEmail: smvahsan@gmail.com",
"article": 9
}
],
"views": 880,
"downloads": 218,
"references": [
{
"id": 231,
"serial_number": 1,
"reference": "Min D, Li Z, et al. The co-regulation of ethylene biosynthesis and ascorbate–glutathione cycle by methy jasmonate contributes to aroma formation of tomato fruit during postharvest ripening. Journal of Agricultural and Food Chemistry. 2020;68:10822-32.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 232,
"serial_number": 2,
"reference": "Huang R, Cheng Y, et al. Postharvest application of acibenzolar- s-methyl delays the senescence of pears by mediating the ascorbate-glutathione cycle. Scientia Horticulturae. 2022;293:110741.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 233,
"serial_number": 3,
"reference": "Iñiguez-Moreno M, González-González RB, et al. Nano and technological frontiers as a sustainable platform for postharvest preservation of berry fruits. Foods. 2023;12:3159.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 234,
"serial_number": 4,
"reference": "Palumbo M, Attolico G, et al. Emerging postharvest technologies to enhance the shelf-life of fruit and vegetables: An overview. Foods. 2022;11:3925.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 235,
"serial_number": 5,
"reference": "Rodrigues M, Ordoñez-Trejo EJ, et al. Dissecting postharvest chilling injuries in pome and stone fruit through integrated omics. Frontiers in Plant Science. 2024;14.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 236,
"serial_number": 6,
"reference": "Zhang W, Jiang Y, et al. The role of different natural organic acids in postharvest fruit quality management and its mechanism. Food Frontiers. 2023;4:1127-43.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 237,
"serial_number": 7,
"reference": "Cuéllar-Torres EA, Aguilera-Aguirre S, et al. Molecular aspects revealed by omics technologies related to the defense system activation in fruits in response to elicitors: A review. Horticulturae. 2023;9:558.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 238,
"serial_number": 8,
"reference": "Gong D, Bi Y, et al. Preharvest elicitors spray improves antioxidant activity, alleviates chilling injury, and maintains quality in harvested fruit. Horticulturae. 2022;8:1208.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 239,
"serial_number": 9,
"reference": "Zhou Y, Liu J, et al. Exogenous glutathione maintains the postharvest quality of mango fruit by modulating the ascorbate-glutathione cycle. PeerJ. 2023;11:e15902.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 240,
"serial_number": 10,
"reference": "Li Z, Min D, et al. The roles of slmyc2 in regulating ascorbate-glutathione cycle mediated by methyl jasmonate in postharvest tomato fruits under cold stress. Scientia Horticulturae. 2021;288:110406.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 241,
"serial_number": 11,
"reference": "Zhu L, Yu H, et al. Effect of methyl jasmonate on the quality and antioxidant capacity by modulating ascorbate-glutathione cycle in peach fruit. Scientia Horticulturae. 2022;303:111216.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 242,
"serial_number": 12,
"reference": "Ali S, Nawaz A, et al. Hydrogen sulfide mitigates chilling injury of postharvest banana fruits by regulating γ-aminobutyric acid shunt pathway and ascorbate–glutathione cycle. Frontiers in Plant Science. 2022;13.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 243,
"serial_number": 13,
"reference": "Ali S, Nawaz A, et al. Carboxymethyl cellulose coating maintains quality of harvested aonla fruit by regulating oxidative stress and ascorbate-glutathione cycle. Postharvest Biology and Technology. 2024;207:112621.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 244,
"serial_number": 14,
"reference": "Wu H, Wang F, et al. Carboxymethyl chitosan different durations induces disease resistance of grapefruit by modulating ascorbate-glutathione cycle and cell wall metabolism. Postharvest Biology and Technology. 2024;211:112845.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 245,
"serial_number": 15,
"reference": "Lu X, Zhang H, et al. Effects of ozone treatment on gene profiling involved in asa-gsh cycle in postharvest cantaloupe. Scientia Horticulturae. 2023;312:111843.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 246,
"serial_number": 16,
"reference": "Yao M, Ge W, et al. Exogenous glutathione alleviates chilling injury in postharvest bell pepper by modulating the ascorbate-glutathione (asa-gsh) cycle. Food Chemistry. 2021;352:129458.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 247,
"serial_number": 17,
"reference": "Imahori Y, Bai J. Postharvest management of fruits and vegetables—series ii. Foods. 2024;13:1049.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 248,
"serial_number": 18,
"reference": "Meitha K, Pramesti Y, et al. Reactive oxygen species and antioxidants in postharvest vegetables and fruits. International Journal of Food Science. 2020;2020:8817778.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 249,
"serial_number": 19,
"reference": "Rajput VD, Harish, et al. Recent developments in enzymatic antioxidant defence mechanism in plants with special reference to abiotic stress. Biology. 2021;10:267.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 250,
"serial_number": 20,
"reference": "Hasanuzzaman M, Bhuyan MHMB, et al. Regulation of ascorbate-glutathione pathway in mitigating oxidative damage in plants under abiotic stress. Antioxidants. 2019;8:384.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 251,
"serial_number": 21,
"reference": "Zheng X, Gong M, et al. Metabolism and regulation of ascorbic acid in fruits. Plants. 2022;11:1602.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 252,
"serial_number": 22,
"reference": "Hasanuzzaman M, Nahar K, et al. Glutathione in plants: Biosynthesis and physiological role in environmental stress tolerance. Physiology and Molecular Biology of Plants. 2017;23:249-68.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 253,
"serial_number": 23,
"reference": "Humbal A, Pathak B. Influence of exogenous elicitors on the production of secondary metabolite in plants: A review (“vsi: Secondary metabolites”). Plant Stress. 2023;8:100166.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 254,
"serial_number": 24,
"reference": "Wang B, Bi Y. The role of signal production and transduction in induced resistance of harvested fruits and vegetables. Food Quality and Safety. 2021;5.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 255,
"serial_number": 25,
"reference": "Ji H, Wang J, et al. Meta-analysis of chitosan-mediated effects on plant defense against oxidative stress. Science of The Total Environment. 2022;851:158212.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 256,
"serial_number": 26,
"reference": "Panahirad S, Dadpour M, et al. Applications of carboxymethyl cellulose- and pectin-based active edible coatings in preservation of fruits and vegetables: A review. Trends in Food Science & Technology. 2021;110:663-73.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 257,
"serial_number": 27,
"reference": "Celi GEA, Gratão PL, et al. Physiological and biochemical roles of ascorbic acid on mitigation of abiotic stresses in plants. Plant Physiology and Biochemistry. 2023;202:107970.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 258,
"serial_number": 28,
"reference": "Wu Y, Li X, et al. Erf subfamily transcription factors and their function in plant responses to abiotic stresses. Frontiers in Plant Science. 2022;13.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 259,
"serial_number": 29,
"reference": "Zhou C, Dong W, et al. Γ-aminobutyric acid treatment induced chilling tolerance in postharvest peach fruit by upregulating ascorbic acid and glutathione contents at the molecular level. Frontiers in Plant Science. 2022;13.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 260,
"serial_number": 30,
"reference": "Yang Z, Zhu S, et al. Nitric oxide modulates folate-mediated one-carbon metabolism and mitochondrial energy levels of peaches during cold storage. Frontiers in Nutrition. 2023;10.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 261,
"serial_number": 31,
"reference": "Anwar A, She M, et al. Biological roles of ornithine aminotransferase (oat) in plant stress tolerance: Present progress and future perspectives. International Journal of Molecular Sciences. 2018;19:3681.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 262,
"serial_number": 32,
"reference": "Botondi R, Barone M, et al. A review into the effectiveness of ozone technology for improving the safety and preserving the quality of fresh-cut fruits and vegetables. Foods. 2021;10:748.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 263,
"serial_number": 33,
"reference": "Aslam R, Alam MS, et al. Sanitization potential of ozone and its role in postharvest quality management of fruits and vegetables. Food Engineering Reviews. 2020;12:48-67.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 264,
"serial_number": 34,
"reference": "Vijay Rakesh Reddy S, Sudhakar Rao DV, et al. Role of ozone in post-harvest disinfection and processing of horticultural crops: A review. Ozone: Science & Engineering. 2022;44:127-46.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 265,
"serial_number": 35,
"reference": "Li C, Tao J, et al. Gaseous ozone regulates reactive oxygen species metabolism and ascorbate–glutathione cycle to delay the senescence of fresh-cut red pitaya (selenicereus undatus) fruit. Scientia Horticulturae. 2023;312:111839.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 266,
"serial_number": 36,
"reference": "Cook R, Lupette J, et al. The role of chloroplast membrane lipid metabolism in plant environmental responses. Cells. 2021;10:706.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 267,
"serial_number": 37,
"reference": "Ho T-T, Murthy HN, et al. Methyl jasmonate induced oxidative stress and accumulation of secondary metabolites in plant cell and organ cultures. International Journal of Molecular Sciences. 2020;21:716.",
"pmc": null,
"DOI": null,
"article": 9
},
{
"id": 268,
"serial_number": 38,
"reference": "Ritonga FN, Chen S. Physiological and molecular mechanism involved in cold stress tolerance in plants. Plants. 2020;9:560.",
"pmc": null,
"DOI": null,
"article": 9
}
]
},
{
"id": 7,
"slug": "280-1713330329",
"featured": true,
"slider": false,
"issue": "Vol2 Issue2",
"type": "original_article",
"manuscript_id": "280-1713330329",
"recieved": "2024-04-17",
"revised": null,
"accepted": "2024-05-21",
"published": "2024-06-01",
"pdf_file": "https://plant-trends.bsmiab.org/media/pdf_file/2024/04/280-1713330329.pdf",
"title": "Nitric oxide facilitates the activation of iron acquisition genes in soybean (<span>Glycine max</span> L.) exposed to iron deficiency",
"abstract": "<p><em>Glycine max</em> L.). Fe-deficiency causes chlorosis, growth retardation, low photosynthetic performance, and productivity. Nitric oxide (NO) is known to serve as a signalling molecule in plants, but its role in mitigating Fe-deficiency in soybeans is still vague. Therefore, the study aims to explore physiological and molecular mechanisms associated with Fe-acquisition and homeostasis is highly demandable. In this study, we found Fe-deficiency occurred leaf chlorosis, inhibiting photosynthetic performance and biomass yield in soybean. However, the exogenous supplementation of sodium nitroprusside, a donor of nitric oxide (NO) significantly restored these physiological attributes. The fluorescence intensity of NO indicates that NO-singling induced in response to Fe-deficiency, along with NO induces conversion of Fe<sup>3+</sup> to Fe<sup>2+</sup>, which leads to Fe acquisition and homeostasis mechanism in Strategy I soybean plants. This study further suggests that NO up-regulates the expression candidate genes <em>ZmIRT1</em>, <em>ZmFRO2,</em> and<em> ZmSultr1;3</em> related to Fe-acquisition and homeostasis in soybean plants. These findings might be useful to soybean breeders and farmers for coping with Fe-deficiency in Strategy I soybean and other grains crops.</p>",
"journal_reference": "Plant Trends. 2024; 2(2): 16-23",
"academic_editor": "Md Azizul Haque, PhD; Hajee Mohammad Danesh Science and Technology University, Bangladesh",
"cite_info": "Bagchi R, Diba SF, et al. Nitric oxide facilitates the activation of iron acquisition genes in soybean (Glycine max L.) exposed to iron deficiency. Plant Trends. 2024; 2(2): 16-23.",
"keywords": [
"Nitric-oxide signaling",
"Glycine max",
"Fe-shortage",
"Chlorosis",
"Strategy-I."
],
"DOI": "10.5455/pt.2024.02",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>Iron (Fe) is a microelement crucial for many physiological processes such as photosynthesis, respiration, DNA synthesis, and nitrogen fixation in plants [<a href=\"#r-1\">1</a>]. Fe-limitation causes leaf chlorosis, plant growth inhibition, mineral shortage, and plant productivity loss in plants including legumes [<a href=\"#r-2\">2</a>]. Fe-deficiency limitation is very common in alkaline soils as well as soils possessing high cation exchange capacity [<a href=\"#r-3\">3</a>]. In high pH soils, the deposition of CaCO₃ forms bicarbonate (HCO⁻₃) suffers system due to inhibition and/or poor aeration that may lead to Fe-shortage in plants [<a href=\"#r-4\">4</a>]. In addition, calcareous soil combined with overwatering initiates Fe deficiency in plants [<a href=\"#r-2\">2</a>]. Fe is essential for successful legume-rhizobium symbiosis. Fe is required for leghemoglobin protein to carry oxygen in cells, for nitrogenase enzyme, and for cytochromes-involving electron transport system (ETS) in bacteroids [<a href=\"#r-5\">5</a>]. <br />\r\nPlants acquire Fe from soils through Strategy-I(dicot and non‐graminaceous species) and strategy-II mechanisms (graminaceous species) [<a href=\"#r-6\">6</a>]. These complex processes allow plants for Fe acquisition and homeostasis through a series of physiological and molecular alterations. In the Fe acquisition process, ferric (Fe<sup>3+</sup>) converts into ferrous (Fe<sup>2+</sup>) presence of ferric chelate reductase (FCR) enzyme that is one of the key mechanisms of Strategy-I plants [<a href=\"#r-7\">7</a>]. Another way is to solubilization of Fe in soils by secreting H+ into the rhizosphere [<a href=\"#r-2\">2</a>]. In addition, Fe transporters play a vital role at transcriptional and posttranscriptional levels for uptalking Fe from soils [8]. Iron-regulated transporter1 (IRT1) is a key transporter gene that uptake Fe2+ in Strategy-I plants [<a href=\"#r-4\">4</a>]. The ferric reduction oxidase 2 (FRO2) coordinates IRT1 responsible for high-affinity Fe uptake from soils [<a href=\"#r-8\">8</a>]. <br />\r\nSignaling molecules are involved in regulating the adaptive response of Fe in plants. Nitric oxide (NO) is a gracious molecule that regulates physiological processes and stress tolerance in plants [<a href=\"#r-9\">9</a>]. NO signaling is involved in Fe-deficiency response and homeostasis in plants [<a href=\"#r-10\">10</a>]. Numerous studies have shown that NO is the key marker of Fe-deficiency response that coordinates the expression of genes and transcription factors related to Fe-regulation in several plant species [<a href=\"#r-4\">4</a>, <a href=\"#r-10\">10</a>, <a href=\"#r-11\">11</a>]. In this scenario, the role of NO in recovering iron deficiency and the response of Fe-responsive candidate genes in legume crops could play a significant role in nutritional security. <br />\r\nSoybean (<em>Glycine max</em> L.) is an important legume and an excellent source of food, feed, and promising biofuel. Fe deficiency is a major limitation in high pH calcareous soils, The Fe-shortage problem leads to a decline in soybean yield equivalent economic loss of $820 million per year [<a href=\"#r-12\">12</a>]. Therefore, eco-friendly, and cost-effective strategies and the discovery of molecular alteration associated with Fe acquisition and homeostasis of grain crops are highly demandable. In this context, we have explored mechanistic insights into NO-mediated regulation of key genes associated with Fe-acquisition and homeostasis in soybean plants.</p>"
},
{
"section_number": 2,
"section_title": "MATERIALS AND METHODS",
"body": "<p><strong>Plant cultivation with Fe-deficiency and NO treatment </strong><br />\r\nDisease-free-viable soybean (<em>Glycine max</em> L.) seeds were surface sterilized with 70% ethanol for 1 min, seeds were washed properly using double distilled water, and then seeds were transferred to a germination tray for three days. Subsequently, 3 days later the soybean’s small seedlings were transferred to micro-macro nutrient-containing traditional Hoagland solution [<a href=\"#r-13\">13</a>]. The soybean seedlings were grown with the following combinations: 25 µM FeNaEDTA with all macro-micro element containing media (control); only 0.1 µM FeNaEDTA (−Fe); 0.1 µM FeNaEDTA +sodium nitroprusside (100 µM) as nitric oxide (NO) donor (-Fe+NO); and 100 µM SNP treatment (NO). All plants were maintained in a controlled environment with 60–65% relative humidity (RH), 200 μmolm<sup>−2</sup> s<sup>−1</sup> light intensity, temperature 25 °C, and 14 h light and 10 h dark combination. The treatments were terminated after two weeks. Samples were collected with liquid N2, and then frozen samples were stored at -80°C until further analyses. At least three biological replications were considered for each treatment. </p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Determination of photosynthetic parameters and plant biomass</strong><br />\r\nThe treated plants were dark adapted for 1 h prior to measuring Fv/Fm and SPAD value (leaf greenness). The maximum yield of photosystem II (Fv/Fm) was measured using a portable fluorometer system (PAM 200, Effeltrich, Germany), while SPAD score was recorded using a SPAD meter (Minolta, Japan). Soybean seedlings were dried at 70°C for 72 h and then weighted using an electrical balance (Mettler PM 200, Switzerland). The mean value of three independent seedlings was graphically presented for each treatment.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Measurement of Fe and S in plants</strong><br />\r\nSoybean leaf samples were dried at 70°C for 72 h and then digested using an acid solution (HClO<sub>4</sub>/HNO<sub>3</sub>; 1:3 v/v), the Fe and S concentrations were determined in plant sample using ICP-MS system (Agilent 7700, Santa Clara, CA, USA). A standard curve was fit using a multi-element ICP-standard-solution (ROTI®STAR, Roth, Germany).</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Fluorescent histochemical staining of NO in soybean root cells</strong><br />\r\nThe nitric oxide (NO) signal was measured using a Fluorescence system (CLS-01-00076, Logos Biosystem, South Korea). NO-specific fluorescent probe 4,5-diaminofluorescein diacetate (DAF-2DA) (Sigma-Aldrich, Burlington, MA, USA) was used for fluorescent intensity. In brief, the control and treated soybean root tips were washed with deionized water properly to remove excess nutrients from the root tip surface, excised them abound 2 mm in size, the excised root tips were incubated 10 mM Tris−HCl buffer (pH 6.5) containing 10 μM DAF-2DA at dark condition for 30 min. Subsequently, the root tips were washed trice with diethylpyrocarbonate (DEPC) treated water. The reaction of the NO signal with DAF-2DA was observed with 495 nm excitation and 515 nm emissions.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Expression analysis of key genes related to Fe-acquisition and homeostasis </strong><br />\r\nTotal RNA was isolated from soybean samples using an RNA extraction kit (QIAGEN, Hilden, Germany). Briefly, 0.1 g of plant tissue was homogenized with extraction buffer containing 2M DDT and 1% (v/v) β-ME. The whole procedure associated with RNA extraction, cDNA synthesis, and quantitative real-time polymerase chain reaction (qRT-PCR) was performed according to the method used previously [<a href=\"#r-14\">14</a>]. The ZmActin was used as internal control, and the expression levels of key genes were calculated using the 2−ΔΔCt method [<a href=\"#r-15\">15</a>].</p>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p><strong>Changes in plant morphology, photosynthesis, and plant biomass</strong><br />\r\nSoybean plants showed significant chlorosis in response to Fe-deficiency (<a href=\"#figure1\">Figure 1</a>). As a consequence, nutrient shortage, photosynthesis efficiency, pigment content, and biomass yield significantly decreased in response to Fe-deficiency (<a href=\"#figure2\">Figure 2</a>A-C). Furthermore, photosynthetic parameters including Fv/Fm and SPAD score were significantly lower under Fe-deficiency compared to control plants (<a href=\"#figure2\">Figure 2</a> A, B). The plant biomass significantly declined under Fe-deficiency (<a href=\"#figure2\">Figure 2</a>C). Interestingly, the supplementation of NO significantly restored these limitations after adding to the Fe-deficient condition, suggesting NO is a potent regulator that is involved in recovering Fe-deficiency in soybean plants. </p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"395\" src=\"/media/article_images/2024/25/04/280-1713330329-Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1</strong>. Morphological alteration of soybean plants in response to 25 µM FeNaEDTA with all macro-micro element containing media (control); 0.1 µM FeNaEDTA (−Fe); 0.1 µM FeNaEDTA +sodium nitroprusside (100 µM) as nitric oxide (NO) donor (-Fe+NO); and 100 µM SNP treatment (NO).</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"176\" src=\"/media/article_images/2024/25/04/280-1713330329-Figure2.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 2</strong>. Changes of physiological attributes soybean plants in response to 25 µM FeNaEDTA with all macro-micro element containing media (control); 0.1 µM FeNaEDTA (−Fe); 0.1 µM FeNaEDTA +sodium nitroprusside (100 µM) as nitric oxide (NO) donor (-Fe+NO); and 100 µM SNP treatment (NO). Fv/Fm (A), SPAD score (B), plant biomass yield (C). Data represents ± SD of three independent biological samples. Different letters above on bar columns indicate significant difference at P < 0.05 level. </figcaption>\r\n</figure>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Regulation of Fe and S concentration </strong><br />\r\nFe-deficiency drastically reduced Fe and S concentrations in soybean leaves compared to the control (<a href=\"#figure3\">Figure 3</a>). However, NO supplementation significantly recovered the acquisitions of both elements in soybeans (<a href=\"#figure3\">Figures 3</a>A, B). NO application to the control plants slightly enhanced Fe and S levels, but the increases were not statistically significant compared to the control. The recovery of Fe and S concentrations in response to NO indicates that NO actively enhanced the levels of these elements, which had declined under Fe deficiency.</p>\r\n\r\n<div id=\"figure3\">\r\n<figure class=\"image\"><img alt=\"\" height=\"237\" src=\"/media/article_images/2024/25/04/280-1713330329-Figure3.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 3</strong>. Alteration of iron (Fe) and sulfur (s) content in soybean plants in response to 25 µM FeNaEDTA with all macro-micro element containing media (control); 0.1 µM FeNaEDTA (−Fe); 0.1 µM FeNaEDTA +sodium nitroprusside (100 µM) as nitric oxide (NO) donor (-Fe+NO); and 100 µM SNP treatment (NO). Fe content (A), S content (B). Data represents ± SD of three independent biological samples. Different letters above on bar columns indicate significant difference at P < 0.05 level. </figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Detection of NO-signal under Fe-deficiency </strong><br />\r\nThe root tip of the Fe-deficient plant induced NO-signal, while a NO-responsive signal was not observed in control plants (<a href=\"#figure4\">Figure 4</a>). The fluorescence intensity of NO was higher after exogenously supplemented NO in Fe-deficient plants which was significantly higher compared to Fe-deficient and control plants (<a href=\"#figure4\">Figure 4</a>A, B). The fluorescence intensity was found to be induced in NO-treated positive control plants. These results indicate that NO signal is induced in response to exogenous endogenous NO in root tips cells, which indicates NO-involving Fe-adaptive mechanisms in soybean plants. </p>\r\n\r\n<div id=\"figure4\">\r\n<figure class=\"image\"><img alt=\"\" height=\"234\" src=\"/media/article_images/2024/25/04/280-1713330329-Figure4.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 4</strong>. Fluorescence intensity of nitric oxide (NO) in soybean root tips in response to 25 µM FeNaEDTA with all macro-micro element containing media (control); 0.1 µM FeNaEDTA (−Fe); 0.1 µM FeNaEDTA +sodium nitroprusside (100 µM) as nitric oxide (NO) donor (-Fe+NO); and 100 µM SNP treatment (NO). NO intensity shows in soybean root tips under different treatments (A), presentation of fluorescence intensity of NO (B). Data represents ± SD of three independent biological samples. Different letters above on bar columns indicate significant difference at P < 0.05 level. </figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Upregulation of <em>GmIRT1, GmFRO2 </em>and <em>GmSultr1;3</em></strong><br />\r\nSeveral key genes related to Fe regulation and homeostasis were induced in response to Fe-deficiency (<a href=\"#figure5\">Figure 5</a>).The expression of <em>GmIRT1</em> was significantly induced by Fe-deficiency that was, even more, higher after exogenous supplementation of NO in plants (<a href=\"#figure5\">Figure 5A</a>). Expression of low-Fe-inducible ferric chelate reductase gene <em>GmFRO2</em> drastically upregulated by Fe-deficiency, indicating Fe-deficiency is significantly reduced at root cells (<a href=\"#figure5\">Figure 5B</a>). In addition, a significant up-regulation pattern was observed for sulfur transporter gene <em>GmSultr1;3</em> compared to control plants (<a href=\"#figure5\">Figure 5B</a>). Hence, the expression of this gene was substantially expressed in response to NO-supplementation in soybean plants (<a href=\"#figure5\">Figure 5C</a>).</p>\r\n\r\n<div id=\"figure5\">\r\n<figure class=\"image\"><img alt=\"\" height=\"176\" src=\"/media/article_images/2024/25/04/280-1713330329-Figure5.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 5</strong>. Expression of key genes involved in Fe-acquisition and homeostasis in soybean in response to 25 µM FeNaEDTA with all macro-micro element containing media (control); 0.1 µM FeNaEDTA (−Fe); 0.1 µM FeNaEDTA +sodium nitroprusside (100 µM) as nitric oxide (NO) donor (-Fe+NO); and 100 µM SNP treatment (NO). ZmIRT1 (A), ZmFRO2 (B), and ZmSultr1;3 (C). Data represents ± SD of three independent biological samples. Different letters above on bar columns indicate significant difference at P <0.05 level.</figcaption>\r\n</figure>\r\n</div>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p>Fe-deficiency-induced leaf chlorosis is obvious in dicot plants that cause growth inhibition, reduction of Fe concentration, and plant productivity [<a href=\"#r-2\">2</a>]. In this study, we have explored mechanistic insights of NO-mediated restoration of these attributes in soybean plants. In the following sections, we discussed the role of NO in regulating physiological and molecular mechanisms, upregulation of key genes related to Fe-acquisition and homeostasis in soybean with other Strategy I plants, and their interpretations.<br />\r\nRestoration of Fe, photosynthetic performance, and plant productivity in soybeans is a promising sign for Strategy I grain crops. NO plays a significant role in reactivating the process related to Fe-acquisition, enhancement of chlorophyll content, and regulation of key genes involved in Fe-regulation in plants [<a href=\"#r-10\">10</a>]. Fe chlorosis and Fe resupply significantly regulate leaf xylem architecture, water relations, gas exchange, and stomatal performance of field-grown peach, which alters its original form and performance of photosystem II (PS-II) and influences the Fe-dependent chlorophyll biosynthesis process [<a href=\"#r-16\">16</a>]. Another study also documented that Fe-deficiency-induced chlorotic leaf negatively regulated PS-II in plants [<a href=\"#r-17\">17</a>]. As a consequence of these events, energy production, and biomass yield are hampered in plants. In this NO-treated soybean, the PS-II (Fv/Fm) rate significantly increased which suggests NO can overcome PS-II damage along with leaf greenness. In addition, NO signal found Fe-deficient roots, but root architecture remains unchanged and/or little improved in NO-treated plants, indicating Fe-deficiency not significantly damaged the root cells compared to the leaf in Strategy I soybean. Our findings suggest that NO can enhance Fe-acquisition and homeostasis processes in soybeans, and NO improves PS-II performance allowing them to translocate sufficient Fe content in the PS-II reaction centre in soybean plants. <br />\r\nPlants have evolved an adaptive response to Fe-deficiency, but this response depends on Strategy I and II mechanisms and plant species. NO restored Fe concentration in the Strategy I alfalfa legume [<a href=\"#r-11\">11</a>]. Moreover, S directly interacts with Fe, and the capacity to uptake Fe is highly dependable on S availability in monocot and dicot plant species [<a href=\"#r-18\">18</a>]. In our current study, the fashion of accumulation patterns of Fe and S are similar suggesting a correlation and common mechanism in Strategy I soybean plants. Hence, this accumulation was enhanced after supplementation of NO in Fe-deficient plants. The overall findings imply that NO supplementation induces Fe and S availability in soybeans and helps to restore the Fe-deficiency problem in soybeans.<br />\r\nThe <em>FRO2</em> gene encodes low-iron-inducible ferric chelate reductase enzyme responsible for the reduction of Fe status in plants, and <em>FRO2</em> coordinates IRT1 that helps to Fe-uptake in <em>Arabidopsis </em>[<a href=\"#r-8\">8</a>]. In this NO-treated soybean, the expression <em>ZmFRO2</em> was highly expressed following the imposition of Fe-starvation plants, while <em>ZmIRT1</em> and <em>ZmSultr1;3</em> were highly induced in response to exogenous supplementation of NO supply. As a result, we noticed that NO may promote the conversion of Fe<sup>3+</sup> to Fe<sup>2+</sup>, which leads to Fe acquisition and homeostasis procession in Strategy I soybean plants. This finding is supported by the observation of the Strategy-I plant where NO played as an inducer of Strategy-I responses to increase Fe availability and mobilization in Fe-starved broccoli [<a href=\"#r-4\">4</a>]. Therefore, the above findings imply that NO-triggers the expression of key genes related to Fe acquisition and homeostasis in soybean plants.</p>"
},
{
"section_number": 5,
"section_title": "CONCLUSION",
"body": "<p>This study implies the mechanistic basis of NO-mediated protection of Fe-deficiency in Strategy soybean plants (<a href=\"#figure6\">Figure 6</a>). Fe-shortage induced chlorosis, declined chlorophyll content, photosynthetic performance, and plant biomass yield in soybean. However, the exogenous supplementation of NO significantly restored these physiological attributes in soybeans. This study further suggests that NO played a potential inducer of key genes (<em>ZmIRT1,</em> <em>ZmFRO2</em>, and<em> ZmSultr1;3</em>) related to Fe-acquisition and homeostasis in Strategy I soybean plants. Additionally, NO helps to induce the conversion of Fe<sup>3+</sup> to Fe<sup>2+</sup>, which leads to Fe acquisition and homeostasis mechanism in Strategy I soybean plants (<a href=\"#figure6\">Figure 6</a>). These findings encourage us to improve Strategy I soybean and other grains crops with Fe-sufficiency through breeding programs.</p>\r\n\r\n<div id=\"figure6\">\r\n<figure class=\"image\"><img alt=\"\" height=\"299\" src=\"/media/article_images/2024/25/04/280-1713330329-Figure6.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 6</strong>. Mechanistic overview of nitric oxide (NO) involving Fe-acquisition and homeostasis in soybean. The NO played a key frontier in inducing candidate genes <em>ZmIRT1, ZmFRO2</em>, and <em>ZmSultr1;3.</em> The <em>ZmIRT1</em> is involved in the conversion of ferric (Fe<sup>3+</sup>) into ferrous (Fe<sup>2+</sup>) in root cells which further leads to Fe-acquisition and homeostasis in soybean. On the other side, NO enhances the accumulation of sulfur (S) through the sulfate transporter <em>ZmSultr1;3.</em> The whole system combinedly enhanced Fe-acquisition and homeostasis in Strategy I soybean plants. </figcaption>\r\n</figure>\r\n</div>"
},
{
"section_number": 6,
"section_title": "ACKNOWLEDGEMENT",
"body": "<p>We thank the Grassland and Forage Division, National Institute of Animal Science, Rural Development, Administration, Cheonan 31000, Republic of Korea for technical support. </p>"
},
{
"section_number": 7,
"section_title": "AUTHOR CONTRIBUTIONS",
"body": "<p>RB performed plant cultivation, processed samples, and measured phenotypic data. SFD collaborated with the expression analysis of genes. SAS and MNA performed physiological observations of plants. MAR performed ICP-MS and microscopic analysis. AHK initiated the research idea and coordinated the whole work. </p>"
},
{
"section_number": 8,
"section_title": "CONFLICTS OF INTEREST",
"body": "<p>There is no conflict of interest among the authors.</p>"
}
],
"figures": [
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/25/04/280-1713330329-Figure1.jpg",
"caption": "Figure 1. Morphological alteration of soybean plants in response to 25 µM FeNaEDTA with all macro-micro element containing media (control); 0.1 µM FeNaEDTA (−Fe); 0.1 µM FeNaEDTA +sodium nitroprusside (100 µM) as nitric oxide (NO) donor (-Fe+NO); and 100 µM SNP treatment (NO).",
"featured": false
},
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/25/04/280-1713330329-Figure2.jpg",
"caption": "Figure 2. Changes of physiological attributes soybean plants in response to 25 µM FeNaEDTA with all macro-micro element containing media (control); 0.1 µM FeNaEDTA (−Fe); 0.1 µM FeNaEDTA +sodium nitroprusside (100 µM) as nitric oxide (NO) donor (-Fe+NO); and 100 µM SNP treatment (NO). Fv/Fm (A), SPAD score (B), plant biomass yield (C). Data represents ± SD of three independent biological samples. Different letters above on bar columns indicate significant difference at P < 0.05 level.",
"featured": false
},
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/25/04/280-1713330329-Figure3.jpg",
"caption": "Figure 3. Alteration of iron (Fe) and sulfur (s) content in soybean plants in response to 25 µM FeNaEDTA with all macro-micro element containing media (control); 0.1 µM FeNaEDTA (−Fe); 0.1 µM FeNaEDTA +sodium nitroprusside (100 µM) as nitric oxide (NO) donor (-Fe+NO); and 100 µM SNP treatment (NO). Fe content (A), S content (B). Data represents ± SD of three independent biological samples. Different letters above on bar columns indicate significant difference at P < 0.05 level.",
"featured": false
},
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/25/04/280-1713330329-Figure4.jpg",
"caption": "Figure 4. Fluorescence intensity of nitric oxide (NO) in soybean root tips in response to 25 µM FeNaEDTA with all macro-micro element containing media (control); 0.1 µM FeNaEDTA (−Fe); 0.1 µM FeNaEDTA +sodium nitroprusside (100 µM) as nitric oxide (NO) donor (-Fe+NO); and 100 µM SNP treatment (NO). NO intensity shows in soybean root tips under different treatments (A), presentation of fluorescence intensity of NO (B). Data represents ± SD of three independent biological samples. Different letters above on bar columns indicate significant difference at P < 0.05 level.",
"featured": true
},
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/25/04/280-1713330329-Figure5.jpg",
"caption": "Figure 5. Expression of key genes involved in Fe-acquisition and homeostasis in soybean in response to 25 µM FeNaEDTA with all macro-micro element containing media (control); 0.1 µM FeNaEDTA (−Fe); 0.1 µM FeNaEDTA +sodium nitroprusside (100 µM) as nitric oxide (NO) donor (-Fe+NO); and 100 µM SNP treatment (NO). ZmIRT1 (A), ZmFRO2 (B), and ZmSultr1;3 (C). Data represents ± SD of three independent biological samples. Different letters above on bar columns indicate significant difference at P <0.05 level.",
"featured": false
},
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/25/04/280-1713330329-Figure6.jpg",
"caption": "Figure 6. Mechanistic overview of nitric oxide (NO) involving Fe-acquisition and homeostasis in soybean. The NO played a key frontier in inducing candidate genes ZmIRT1, ZmFRO2, and ZmSultr1;3. The ZmIRT1 is involved in the conversion of ferric (Fe3+) into ferrous (Fe2+) in root cells which further leads to Fe-acquisition and homeostasis in soybean. On the other side, NO enhances the accumulation of sulfur (S) through the sulfate transporter ZmSultr1;3. The whole system combinedly enhanced Fe-acquisition and homeostasis in Strategy I soybean plants.",
"featured": false
}
],
"authors": [
{
"id": 19,
"affiliation": [
{
"affiliation": "Department of Botany, University of Rajshahi, Rajshahi 6205, Bangladesh"
}
],
"first_name": "Ruby",
"family_name": "Bagchi",
"email": null,
"author_order": 1,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 7
},
{
"id": 20,
"affiliation": [
{
"affiliation": "International Rice Research Institute (IRRI), Bangladesh Office, Dhaka, Bangladesh"
}
],
"first_name": "Sheikh Farah",
"family_name": "Diba",
"email": null,
"author_order": 2,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 7
},
{
"id": 21,
"affiliation": [
{
"affiliation": "Department of Botany, University of Rajshahi, Rajshahi 6205, Bangladesh"
}
],
"first_name": "Shifat Ara",
"family_name": "Saiful",
"email": null,
"author_order": 3,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 7
},
{
"id": 22,
"affiliation": [
{
"affiliation": "Department of Botany, University of Rajshahi, Rajshahi 6205, Bangladesh"
}
],
"first_name": "Mst Nahida",
"family_name": "Akter",
"email": null,
"author_order": 4,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 7
},
{
"id": 23,
"affiliation": [
{
"affiliation": "Grassland and Forages Division, Rural Development Administration, Cheonan, South-Korea"
},
{
"affiliation": "ABEx Bio-Research Center, Dhaka, Bangladesh"
}
],
"first_name": "Md Atikur",
"family_name": "Rahman",
"email": "pteditorialoffice@gmail.com",
"author_order": 5,
"ORCID": "https://orcid.org/0000-0001-6779-9599",
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 7
},
{
"id": 24,
"affiliation": [
{
"affiliation": "School of Sciences, University of Louisiana Monroe, Monroe, USA"
}
],
"first_name": "Ahmad Humayan",
"family_name": "Kabir",
"email": "kabir@ulm.edu",
"author_order": 6,
"ORCID": "https://orcid.org/0000-0001-6893-9418",
"co_first_author": false,
"co_author": false,
"corresponding": true,
"corresponding_author_info": "Ahmad Humayan Kabir, PhD; School of Sciences, University of Louisiana at Monroe, Monroe, LA 71209, USA. Email: kabir@ulm.edu",
"article": 7
}
],
"views": 1370,
"downloads": 288,
"references": [
{
"id": 213,
"serial_number": 1,
"reference": "He X-L, Zhang W-Q, et al. Hydrogen sulfide and nitric oxide regulate the adaptation to iron deficiency by affecting Fe homeostasis and thiol redox modification in Glycine max seedlings. Plant Physiol Biochem. 2023;194:1-14.",
"pmc": null,
"DOI": null,
"article": 7
},
{
"id": 214,
"serial_number": 2,
"reference": "Kabir AH, Rahman MA, et al. Mechanistic assessment of tolerance to iron deficiency mediated by Trichoderma harzianum in soybean roots. J Appli Microbiol. 2022;133:2760-78.",
"pmc": null,
"DOI": null,
"article": 7
},
{
"id": 215,
"serial_number": 3,
"reference": "Rahman MA, Bagchi R, et al. Physiological and molecular characterization of strategy-I responses and expression of Fe-transporters in Fe-deficient soybean. S Afr J Bot. 2022;147:942-50.",
"pmc": null,
"DOI": null,
"article": 7
},
{
"id": 216,
"serial_number": 4,
"reference": "Kabir AH, Ela EJ, et al. Nitric oxide acts as an inducer of Strategy-I responses to increase Fe availability and mobilization in Fe-starved broccoli (Brassica oleracea var. oleracea). Plant Physiol Biochem. 2023;194:182-92.",
"pmc": null,
"DOI": null,
"article": 7
},
{
"id": 217,
"serial_number": 5,
"reference": "Brear EM, Day DA, et al. Iron: an essential micronutrient for the legume-rhizobium symbiosis. Front Plant Sci. 2013;4:359.",
"pmc": null,
"DOI": null,
"article": 7
},
{
"id": 218,
"serial_number": 6,
"reference": "Aznar A, Chen NWG, et al. Immunity to plant pathogens and iron homeostasis. Plant Sci. 2015;240:90-7.",
"pmc": null,
"DOI": null,
"article": 7
},
{
"id": 219,
"serial_number": 7,
"reference": "Rahman MA, Bagchi R, et al. Physiological and molecular characterization of strategy-I responses and expression of Fe-transporters in Fe-deficient soybean. S Afr J Bot. 2022;147:942-50.",
"pmc": null,
"DOI": null,
"article": 7
},
{
"id": 220,
"serial_number": 8,
"reference": "Connolly EL, Campbell NH, et al. Overexpression of the FRO2 ferric chelate reductase confers tolerance to growth on low iron and uncovers posttranscriptional control. Plant Physiol. 2003;133:1102-10.",
"pmc": null,
"DOI": null,
"article": 7
},
{
"id": 221,
"serial_number": 9,
"reference": "Lee K-W, Rahman MA, et al. Nitric oxide-induced proteomic analysis in rice leaves. Plant Biotechnol Rep. 2019;13:375-87.",
"pmc": null,
"DOI": null,
"article": 7
},
{
"id": 222,
"serial_number": 10,
"reference": "Zhai L, Xiao D, et al. Nitric oxide signaling is involved in the response to iron deficiency in the woody plant Malus xiaojinensis. Plant Physiol Biochem. 2016;109:515-24.",
"pmc": null,
"DOI": null,
"article": 7
},
{
"id": 223,
"serial_number": 11,
"reference": "Rahman MA, Kabir AH, et al. Nitric oxide prevents Fe deficiency-induced photosynthetic disturbance and oxidative stress in alfalfa by regulating Fe acquisition and antioxidant defense. Antioxidants 2021;10:1556.",
"pmc": null,
"DOI": null,
"article": 7
},
{
"id": 224,
"serial_number": 12,
"reference": "Hansen NC, Jolley VD, et al. Iron deficiency of soybean in the North Central U.S. and associated soil properties. Soil Sci Plant Nutr. 2004;50:983-7.",
"pmc": null,
"DOI": null,
"article": 7
},
{
"id": 225,
"serial_number": 13,
"reference": "Hoagland DR, Arnon DI. The water-culture method for growing plants without soil. Circular Cali Agric Exp Stat. 1950;347:32.",
"pmc": null,
"DOI": null,
"article": 7
},
{
"id": 226,
"serial_number": 14,
"reference": "Rahman MA, Song Y, et al. Mechanistic basis of silicon mediated cold stress tolerance in alfalfa (Medicago sativa L.). Silicon. 2023.",
"pmc": null,
"DOI": null,
"article": 7
},
{
"id": 227,
"serial_number": 15,
"reference": "Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods. 2001;25:402-8.",
"pmc": null,
"DOI": null,
"article": 7
},
{
"id": 228,
"serial_number": 16,
"reference": "Petrou K, Trimborn S, et al. The impact of iron limitation on the physiology of the Antarctic diatom Chaetoceros simplex. Mar Biol. 2014;161:925-37.",
"pmc": null,
"DOI": null,
"article": 7
},
{
"id": 229,
"serial_number": 17,
"reference": "Eichert T, Peguero-Pina JJ, et al. Effects of iron chlorosis and iron resupply on leaf xylem architecture, water relations, gas exchange and stomatal performance of field-grown peach (Prunus persica). Physiol Plant. 2010;138:48-59.",
"pmc": null,
"DOI": null,
"article": 7
},
{
"id": 230,
"serial_number": 18,
"reference": "Astolfi S, Celletti S, et al. Interaction Between Sulfur and Iron in Plants. Frontiers in plant science. 2021;12:670308.",
"pmc": null,
"DOI": null,
"article": 7
}
]
},
{
"id": 6,
"slug": "280-1708590809",
"featured": false,
"slider": false,
"issue": "Vol2 Issue1",
"type": "original_article",
"manuscript_id": "280-1708590809",
"recieved": "2024-02-26",
"revised": null,
"accepted": "2024-04-07",
"published": "2024-04-15",
"pdf_file": "https://plant-trends.bsmiab.org/media/pdf_file/2024/01/280-1708590809.pdf",
"title": "Screening of phytochemicals, antioxidant activity, and in vivo safety profile of the hydroethanolic peel extract of <span>Musa sapientum</span>",
"abstract": "<p>Banana peels can be utilized in many ways such as nutraceuticals to prevent or cure diseases. Therefore, it is important to understand the phytochemical composition, antioxidant capacity, and safety profile of <em>Musa sapientum</em> variety Muraru peels. The study identified the chemical composition of unripe and ripe banana (Muraru) peels of the <em>Musa sapientum</em>, along with their antioxidant capacity, and evaluated the oral acute and sub-acute toxicity of the hydroethanolic extracts. Qualitative and quantitative phytochemical analyses were performed, and GC-MS was used in the identification of bioactive compounds present. The DPPH total antioxidant activity was used to evaluate the scavenging percentage and IC<sub>50</sub>. Acute and sub-acute toxicity tests were done for the hydroethanolic peel extract. Haematology, biochemistry, and relative organ weight were analyzed, and the sub-acute group organs were further analyzed for histopathology. Flavonoids, tannins, and phenols were found abundant in both unripe and ripe banana peels. GC-MS showed that the peels were rich with lipids, fatty acids, and terpenoids. The unripe peels showed significant total antioxidant capacity. No significant difference (p> 0.05) was observed in haematology, biochemistry, or relative organ weight compared to the normal controls. Histopathological examination of the liver, lung, heart, kidney, and spleen showed normal tissue orientation in all treated groups. Unripe and ripe banana peels both contained rich secondary metabolites, which greatly influence their antioxidant capacity. The toxicity profile indicated that the hydroethanolic peel extract is nontoxic.</p>",
"journal_reference": "Plant Trends. 2024; 2(1): 01-15",
"academic_editor": "Md Jamal Uddin, PhD; ABEx Bio-Research Center, Dhaka, Bangladesh",
"cite_info": "Mahora MC, Kigundu A, et al. Screening of phytochemicals, antioxidant activity, and in vivo safety profile of the hydroethanolic peel extract of Musa sapientum. Plant Trends. 2024; 2(1): 01-15.",
"keywords": [
"Secondary metabolites",
"Bioactive compounds",
"Oxidative stress",
"Nutraceuticals",
"Banana peels"
],
"DOI": "10.5455/pt.2024.01",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>Banana fruit plays a crucial role in human nutrition around the world, and it is good for digestive and heart health. Consuming bananas helps boost potassium, magnesium, vitamin B6, and they are rich in serotine and dopamine, which are neurotransmitters that help improve mood and reduce stress. The plant belongs to the Musaceae family, genus Musa, this fruit category encompasses around 70–75 species globally [<a href=\"#r-1\">1</a>], which includes over 1,000 distinct banana varieties [<a href=\"#r-2\">2</a>]. Remarkably, only a handful of these varieties gain recognition in health circles. Notably, Musa sapientum variety Muraru (local name in Maragua, Kenya), unlike the familiar golden yellow color bananas, it remains green or green-yellow when ripe [<a href=\"#r-3\">3</a>]. Additionally, Muraru banana exhibit thicker peels and a firmer texture compared to Pisang Mas [<a href=\"#r-3\">3</a>]. Although Onyango et al. [<a href=\"#r-3\">3</a>] acknowledged its similarities to Cavendish and Gros Michel, Muraru peels remain unexplored in terms of the potential that could provide additional benefits to human health. <br />\r\nResearch has shown that banana peels have a higher abundance of bioactive compounds compared to the pulp [<a href=\"#r-4\">4</a>]. Phytochemicals are natural compounds found in plants that have various biological effects on the human body. A previous study showed that banana peels are rich in phenolic compounds, carotenoids, flavonoids, tannins, terpenes, alkaloids, glycosides, and phytosterols, all of which offer significant dietary benefits with positive effects on human health and well-being [<a href=\"#r-5\">5</a>]. Endogenous antioxidants are molecules produced naturally within the body that help neutralize harmful free radicals and protect cells from oxidative damage. Exogenous antioxidants help fix this imbalance by lowering free radicals, promoting the growth of healthy cells, protecting cells from premature and abnormal ageing, fighting age-related molecular degeneration, and boosting the immune system, which prevents or manages chronic diseases [<a href=\"#r-6\">6</a>]. Bioactive compounds have various mechanisms and properties to function as antioxidants, anti-inflammatory, anticancer, and antidiabetic agents and strengthen the body's defenses against various illnesses [<a href=\"#r-5\">5</a>].<br />\r\nHowever, bioactive compounds classified as secondary metabolites, while known for their pharmacological effects, can also exhibit toxicological properties in both humans and animals [<a href=\"#r-7\">7-9</a>]. It is important to evaluate the toxicological properties of medicinal plants before using them to make new drugs or improve the effectiveness of treatments that are already in use [<a href=\"#r-7\">7</a>]. Studies have demonstrated that non-nutritive parts of the banana plant, such as the flower, pseudo-stem, and leaf, are non-toxic [<a href=\"#r-10\">10</a>]. Nevertheless, the toxicity profile of banana peels remains inadequately elucidated. Therefore, it is imperative to elucidate the safety profile of Muraru peels.<br />\r\nThe abundant presence of bioactive compounds in banana peels positions them as promising candidates for nutraceutical applications. However, it is crucial to elucidate both the antioxidant capacity and toxicological profile of banana peels. This study aims to assess the acute and subacute toxicity of Muraru peel hydro-ethanol extract, as well as its antioxidant activity and phytoconstituents.</p>"
},
{
"section_number": 2,
"section_title": "MATERIALS AND METHODS",
"body": "<p><strong>Plant material and preparation</strong><br />\r\n<em>M. sapientum </em>variety ‘Muraru’ bananas were bought from a local farm in Maragua, Kenya. Verification of the banana species was done at the Jomo Kenyatta University of Agriculture and Technology (JKUAT) Botany Department, and the herbarium accession number is MCM- JKUATBH/001/2023. The bananas were acquired at their mature unripe stage, and unripe and ripe stages 1 and 4, respectively [<a href=\"#r-11\">11</a>], where used in this study. The peels were prepared for lyophilization according to [<a href=\"#r-12\">12</a>]. After lyophilization, peels were blended into powder and stored in zip-lock bags wrapped with aluminum foil in a -20<sup>0</sup>C refrigerator for further studies. </p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Sample extraction</strong><br />\r\nThe cold maceration extraction method was employed for both unripe and ripe peels using two different solvents: absolute methanol and 70% ethanol (hydro-ethanolic), following the procedures outlined by Oyeyinka and Afolayan [<a href=\"#r-13\">13</a>] and Siddique et al. [<a href=\"#r-14\">14</a>]. The ratio of solid to solvent used was 1:10, and the mixtures were soaked for 72 hours at 21°C in a shaker rotating at 121 rpm. Afterward, they were filtered using a 0.1 Whatman filter paper.</p>\r\n\r\n<p><br />\r\n<strong>Phytochemical characterization and screening</strong><br />\r\n<em>Qualitative phytochemical screening</em><br />\r\nTo identify the presence of phytochemical derivatives in methanolic and hydroethanolic extracts, standard phytochemical screening was conducted. The detection of flavonoids, tannins, saponins, steroids, alkaloids, glycosides, phytosterols, phenols, and terpenoids was detected according to the protocol by Kibria & Kar [<a href=\"#r-15\">15</a>]. </p>\r\n\r\n<p> </p>\r\n\r\n<p><em>Quantitative phytochemical screening</em><br />\r\nThe quantitative phytochemical analysis of total phenolic content (TPC), total tannin content (TTC), and total flavonoid content (TFC) were determined according to the standard protocols of Oyeyinka and Afolayan [<a href=\"#r-13\">13</a>], Aboul-enein et al. [<a href=\"#r-16\">16</a>], and Santhosh and Suriyanarayanan [<a href=\"#r-17\">17</a>]. </p>\r\n\r\n<p> </p>\r\n\r\n<p><em>Gas chromatography mass spectrometry </em><br />\r\nThe Shimadzu GC-MS (Qp2010SE, Japan) was utilized, and the National Institute of Standards and Technology (NIST) library was employed to predict the names of scanned compounds based on the mass-to-charge ratios of the compounds.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Total antioxidant capacity </strong><br />\r\nThe DPPH (1,1-diphenyl-2-picrylhydrazyl) assay was conducted according to Chaudhary et al. [<a href=\"#r-18\">18</a>]. The absorbance was measured at 517 nm using a spectrophotometer (UV 1800, Shimadzu).<br />\r\nScavenging ability (%)= Abs(control) – Abs(sample)/Abs(control) x 100 <br />\r\nwhere Abs (control) indicates the absorbance of DPPH radical + methanol, and Abs (sample) is the absorbance of DPPH radical + sample extract or standard.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Experimental animals</strong><br />\r\nAlbino rats of the Wistar strain, weighing between 150 to 190 g and aged between 7-8 weeks, were obtained from the Small Animal Facility for Research and Innovation (SAFARI) of the Jomo Kenyatta University of Science and Technology (JKUAT). Twenty-one female animals were housed under standard laboratory conditions of 22 ± 3 °C, a relative humidity of 30%, and a 12 h light and dark cycle. The rats had free access to a standard pellet diet (Unga Feeds™) and water ad libitum. Animal bedding, comprising shredded paper, was changed twice a week. <br />\r\nIn accordance with ethical standards, the Animal Use and Care Ethics Review Committee at Jomo Kenyatta University of Science, Agriculture, and Technology, Kenya (JKU/ISERC/02316/1014) granted ethical approval. The conducted experiments were designed to minimize animal suffering, and the experimental protocols strictly followed the guidelines outlined in the National Institutes of Health’s Guide for the Care and Use of Laboratory Animals [<a href=\"#r-19\">19</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Analysis of acute toxicity </strong><br />\r\nThe acute toxicity of the hydroethanolic extract was tested to determine the safety of the agent following the guidelines set by the Organization for Economic Cooperation and Development (OECD) No. 423 [<a href=\"#r-20\">20</a>]. Twelve healthy females in four groups (n = 3) were used, receiving 2000 mg/kg, 1000 mg/kg, 500 mg/kg, and the normal control group received 0.5% Tween 80 dissolved in distilled water, used as a vehicle, with a dissolving volume of 1 ml/100 g. The animals were allowed to acclimatize for one week before experimentation. Prior to the toxicity test, animals were fasted overnight but provided with water. After oral administration of the hydroethanolic peel extract, with the use of oral gavage, animals were observed for the first 30 min, 4 h, and 24 h for signs of morbidity, mortality and once daily over a period of 14 days [<a href=\"#r-20\">20-21</a>]. On the 15th day, all the animals were fasted for 12 h and then euthanized using chloroform. No sign of morbidity or mortality was observed, therefore, the approximate lethal dose (LD<sub>50</sub>) of the peel hydroethanolic extract was estimated to be higher than 2000 mg/kg.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Analysis of subacute toxicity </strong><br />\r\nThe subacute toxicity analysis of the hydroethanolic extract was assessed on nine healthy female animals (n = 3). The normal control group received Tween 80 (0.5%) dissolved in distilled water; two groups received 500 mg/kg and 250 mg/kg of the hydroethanolic peel extract, dissolved in 1 m/100 g, administered orally once every day for a period of 28 days. Animals were observed daily for any sign of morbidity. On the 29th day, all the animals were fasted for 12 h and then euthanized [<a href=\"#r-20\">20</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Preclinical evaluation</strong><br />\r\nThe general behavior, body weight, and feed-water intake of the rats were observed during the acclimatization period. After the administration of the hydroethanolic peel extract, each rat was continuously monitored [<a href=\"#r-22\">22</a>]. </p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Hematology analysis </strong><br />\r\nBlood samples were obtained via cardiac puncture using a syringe and needle, and they were deposited into EDTA-containing tubes. Hematological parameters were conducted using the automated Mindray BC-5000 hematological analyzer from Shenzhen, China [<a href=\"#r-23\">23</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Biochemical analysis</strong><br />\r\nBlood samples for biochemical analysis were placed into non-anticoagulant tubes for sera. An automated Roche-Reflotron dry chemistry analyzer (Roche Diagnostics, Germany) was employed for liver and kidney damage markers, alanine transaminase (ALT), aspartate transaminase (AST), creatinine, and urea, respectively [<a href=\"#r-24\">24</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Histopathology analysis</strong><br />\r\nAfter euthanasia, the liver, lung, kidney, spleen, and heart organs were surgically removed, examined for abnormalities, and weighed. Relative organ weight (ROW) was calculated using the following equation as described [<a href=\"#r-25\">25</a>].<br />\r\nRelative organ weight = Absolute organ weight (g)/ Body weight (g) of rats on final day × 100 <br />\r\nThe subacute-treated group organs were taken for histopathological analysis, according to Kamsu et al. [<a href=\"#r-26\">26</a>]. Stained tissue sections were photographed using photomicrographs and documented for subsequent analysis by a pathologist.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Statistical analyses</strong><br />\r\nAll the data was expressed as mean ± standard error of the mean (SEM). The difference between treated and control groups of acute and subacute toxicity tests were determined by one-way analysis of variance (ANOVA). Values were considered significant at p < 0.05. All statistical analysis was done using GraphPad Prism (student) version 8.4.3 for Windows.</p>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p><strong>Qualitative and quantitative phytochemical profile </strong><br />\r\nThe preliminary qualitative phytochemical profile of Musa sapientum var Muraru showed unripe peels had a significant number of compounds in the hydroethanolic extracts. While the ripe peel showed more compounds in methanolic extracts. Flavonoids, tannins, phytosterols, phenols, and terpenoids secondary metabolites were present across all the peel extracts (<a href=\"#Table-1\">Table 1</a>). The quantitative phytochemical profile showed that unripe and ripe Muraru peels are rich in phenolics, tannins, and flavonoids (the most abundant) (<a href=\"#Table-1\">Table 1</a>). </p>\r\n\r\n<div id=\"Table-1\">\r\n<p><a href=\"https://plant-trends.bsmiab.org/table/280-1708590809-table1/\">Table 1</a><strong>Table 1</strong>. Qualitative and quantitative phytochemical profile for unripe and ripe Muraru peels.</p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Gas chromatography- mass spectrophotometry of Muraru peels </strong><br />\r\nThe chromatograms of unripe and ripe peels confirmed the presence of unsaturated fatty acids, saturated fatty acids, triterpenoids, terpenoids, fatty acids, steroids, and vitamin E (<a href=\"#figure1\">Figure 1</a>). Unripe ethanolic peel extracts revealed seven compounds, with terpenoids (92.82%) being the most abundant. The unripe methanolic extract had five compounds, comprising terpenoids (85.32%) and triterpenoids (12.05%). The ripe ethanolic extract exhibited six compounds, with terpenoids (92.40%) being the most predominant. The ripe methanolic extract contained five compounds, including fatty acids (31.87%), unsaturated fatty acids (30.88%), and lipids (30.22%).</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"474\" src=\"/media/article_images/2024/58/09/280-1708590809-Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1</strong>. GC-MS chromatogram of Muraru unripe and ripe peel extracts. A) Unripe ethanolic extracts showing the presence of undecane (alkane), hexadecenoic acid, methyl ester (saturated fatty acid), 9,12-octadecadienoic acid, methyl ester (unsaturated fatty acid), methyl 8,11,14-heptadecatrienoate (fatty acid), thunbergol (triterpenoid), and 9,19-Cycloergost-24(28)-en-3-ol, 4,14-dimethyl-, acetate, (3β,4α,5α)- (terpenoids). B) Unripe methanolic extracts showing the presence of 2,4-Di-tert-butylphenol (phenols), Pyrrolidine-2-carboxylic acid, methyl-phenyl-amide (saturated heterocycle), Decanedioic acid, bis(2-ethylhexyl) ester (fatty acid), 9,19-Cyclolanostan-3-ol, 24-methylene-, (3β)- (triterpenoid) and 9,19-Cycloergost-24(28)-en-3-ol, 4,14-dimethyl-, acetate, (3β,4α,5α)- (terpenoids). C) Ripe ethanolic extracts showing the presence of melezitose (trisaccharide), Hexadecanoic acid, ethyl ester (saturated fatty acid), 3,7,11,15-Tetramethyl-2-hexadecen-1-OL (phytol), 9,19-Cycloergost-24(28)-en-3-ol, 4,14-dimethyl-, acetate, (3β,4α,5α)- (terpenoids) and Androst-1-en-3-one, 4,4-dimethyl-, (5alpha)- (steroid). D) Ripe methanolic 14-methyl-pentadecanoic acid, methyl ester (fatty acid), 9,12-Octadecadienoic acid, methyl ester (unsaturated fatty acid), 9,12,15-Octadecatrienoic acid, methyl ester (lipid), Heptadecanoic acid, 15-methyl-, methyl ester (saturated fatty acid) and alpha. Tocopheryl acetate (vitamin E). </figcaption>\r\n</figure>\r\n</div>\r\n\r\n<p><br />\r\n<strong>Effect of Muraru peels extract on antioxidant activity</strong><br />\r\nUnripe peel extracts showed significant total antioxidant capacity compared to the ripe peel extracts (<a href=\"#figure2\">Figure 2</a>). Further, the effect of the activity was seen in the Inhibitory Concentration at 50%, where the unripe peel extracts showed a lower IC50 compared to the ripe peel extracts. Unripe methanolic and hydroethanolic extracts have an IC<sub>50</sub> of 73.70 μg/ml; 102.77 μg/ml respectively. While the IC<sub>50</sub> for methanol and hydroethanolic ripe peel extracts were 147.99 μg/ml; 243.45 μg/ml respectively. The lower the IC<sub>50</sub> value, the higher the antioxidant capacity.</p>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"287\" src=\"/media/article_images/2024/58/09/280-1708590809-Figure2.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 2</strong>. The colorimetric DPPH antioxidant assay revealed that Muraru peels have significant antioxidant capacity. Total antioxidant capacity varied with the peel state, with the unripe peels in both solvents revealing the highest scavenging percentage compared to the ripe peel state. The scavenging percentage of the peel extracts against free radical DPPH was 70.12%, 63.99%, 59.91%, and 48.06% at the highest concentration (200 μg/ml) for unripe methanolic, unripe hydroethanolic, ripe peel methanolic, and ripe peel hydroethanolic extracts, respectively. Ascorbic acid was used as the standard for the assay, which also showed that the scavenging percentage increases with an increase in concentration, with 200 μg/mL being the optimum concentration. </figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Effect of Muraru peels extract on acute toxicity</strong><br />\r\nHydroethanolic extracts were used to assess the toxicity of the peels. During the acute toxicity of the peel, no sign of morbidity was noted, and no motility was incurred throughout the doses used in the 14-day period of observation. All animals displayed normal clinical signs and behavior. The weight of the animals during the acute toxicity assessment showed a gradual increase from the initial stage to the end of the treatment (<a href=\"#figure3\">Figure 3</a>A).<br />\r\nHemogram analysis showed that all doses of Muraru peel hydroethanolic extract did not induce significant changes (p > 0.05) in all the measured parameters (<a href=\"#Table-2\">Table 2</a>). Furthermore, there was no alteration in kidney and liver enzymes. Creatinine, urea, ALT, and AST values showed no significant (p > 0.05) changes among the groups (<a href=\"#Table-2\">Table 2</a>).</p>\r\n\r\n<div id=\"figure3\">\r\n<figure class=\"image\"><img alt=\"\" height=\"258\" src=\"/media/article_images/2024/58/09/280-1708590809-Figure3.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 3</strong>. The effect of Muraru peel hydroethanolic extract on body and organ weights during the acute and subacute toxicity tests. A) The mean weight of animals during the acute toxicity study. B) The mean body weight of Wistar rats during the subacute study. C) The relative organ weight obtained after the acute toxicity study. D) The relative organ weight obtained after the subacute toxicity study. The organs in both acute and subacute toxicity tests during harvesting did not show any change in color or enlargement. Relative organ weight was calculated as a percentage of the absolute weight. Values are expressed as a mean ± SEM (standard error mean). The error bars represent SEM (n = 3 rats/group).</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<div id=\"Table-2\">\r\n<p><a href=\"https://plant-trends.bsmiab.org/table/280-1708590809-table2/\">Table 2</a><strong>Table 2</strong>. Effects of Muraru peel ethanol extract on blood parameters in the acute toxicity study. </p>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Effect of Muraru peels extract on subacute toxicity</strong><br />\r\nThe subacute toxicity of the hydroethanolic peel extract showed no sign of morbidity in all the doses, and no mortality occurred throughout the study. There was significant weight gain, which was noticed over the 28 days of subacute toxicity testing in all the groups (<a href=\"#figure3\">Figure 3</a>B). The hemogram assay showed no significant difference (p > 0.05) between the treatment groups and the control group (<a href=\"#Table-3\">Table 3</a>). Serum biochemistry of the liver and kidney enzymes showed no significant difference (p > 0.05) from the control group (<a href=\"#Table-3\">Table 3</a>). The relative organ weight of the treatment groups showed no significant differences (p > 0.05) among the groups (<a href=\"#figure3\">Figure 3</a>D).<br />\r\nThe histopathology of the lung, liver, heart, spleen, and kidney was conducted for the subacute toxicity test, where administration of the hydroethanolic extract was done orally for 28 days. When the histology of the lung, liver, heart, spleen, and kidney from the treated animals was examined, normal morphological features were observed (<a href=\"#figure4\">Figure 4</a>).</p>\r\n\r\n<div id=\"figure4\">\r\n<figure class=\"image\"><img alt=\"\" height=\"627\" src=\"/media/article_images/2024/58/09/280-1708590809-Figure4.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 4</strong>. Morphologies of various organs following sub-acute toxicity. Micrographs of lung, liver, heart, spleen and kidney after 28 days of administration of hydroethanolic Muraru peel extracts. Hematoxylin and eosin stain (Magnification 400×).</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<div id=\"Table-3\">\r\n<p><a href=\"https://plant-trends.bsmiab.org/table/280-1708590809-table3/\">Table 3</a><strong>Table 3</strong>. Effects of Muraru peel hydroethanolic extract on blood parameters in the subacute toxicity.</p>\r\n</div>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p>Over centuries, people have been using plant materials to improve health, treat, and manage various diseases due to the presence of secondary metabolites. This study explored the qualitative phytochemicals of unripe and ripe peels. Both unripe and ripe peels showed the presences of flavonoids, tannins, phytosterols, phenols, alkaloids, and terpenoids. The study results aligned with those by Kibria and Kar [<a href=\"#r-15\">15</a>] and were consistent with research on unripe Kepok and Kultuk peels conducted by Agung et al. [<a href=\"#r-27\">27</a>]. The qualitative results showed the effects of solvent polarity in phytochemical extraction as they varied in number. These results were similar to those of González-Montelongo et al. [<a href=\"#r-28\">28</a>] and Sundaram and Anjum [<a href=\"#r-29\">29</a>]. A study by Ehiowemwenguan et al. [<a href=\"#r-30\">30</a>], showed that ethanol solvent was the best for the extraction of phytochemicals in <em>M. sapientum</em> peels. <br />\r\nThe quantitative profile revealed that both unripe and ripe peels are rich in phenolics, tannins, and flavonoids (most abundant). The findings of this study are consistent with previous studies on the phytochemical composition of banana peels, including Kepok and Kultuk varieties [<a href=\"#r-27\">27</a>], and extraction methods utilizing ethanol solvent [<a href=\"#r-30\">30</a>], notable differences were observed. For instance, the quantitative profile of Muraru peels revealed a higher abundance of flavonoids compared to total tannins, differing from the ratios reported in <em>M. sinensis</em> peels [<a href=\"#r-13\">13</a>]. Additionally, chromatographic analysis highlighted the presence of terpenoids, triterpenoids, lipids, and fatty acids, consistent with prior studies on <em>M. sapientum </em>peels [<a href=\"#r-31\">31-32</a>] albeit with variations in compound diversity.<br />\r\nFurther, the chromatogram of the peels showed an abundance of terpenoids, triterpenoids, lipids, and fatty acids. Puraikalan [<a href=\"#r-32\">32</a>] study revealed that M. sapientum peels were rich in terpenoids and triterpenoids, although their varieties showed more compounds than the current study. The study by Mordi et al. [<a href=\"#r-31\">31</a>] also revealed the presence of fatty acids, palmitic acid, linoleic acids, and methyl esters. However, a difference was noted, as their study showed more compounds than the current study. The extracts showed significant antioxidant activity with unripe peel, with higher activity compared to ripe peel in both methanolic and hydroethanolic extracts. Furthermore, unripe peel extract showed the lowest IC<sub>50</sub> compared to the ripe peels. Mature unripe peel extracts showed increased antioxidant activity compared to mature ripe peel extracts; these results correspond with the study of Sandaram et al. [<a href=\"#r-29\">29</a>]. Note that the current study’s lower IC<sub>50</sub> value indicates high antioxidant activity in the sample. The banana extracts showed strong and moderate antioxidant activity, affirming their efficacy in combating oxidative stress [<a href=\"#r-33\">33</a>]. <br />\r\nSecondary metabolites are produced in nature to serve survival functions for plants and these have also played an important role in the improvement of the health of humans. When comparing the secondary metabolites found in Muraru peels, a variation was seen compared with other studies. This greatly influenced the qualitative, quantitative, GC-MS, and antioxidant capacity of Muraru peels. This is attributed to differences in banana varieties used, the extraction methods employed, the state and quality of the solvent, and variances in geographical locations. Geographical factors such as soil type, climate, altitude, and sunlight exposure can influence the composition of phytochemicals. This diverse array of phytochemicals contributes to the richness and uniqueness of the plant's bioactive compounds. <br />\r\nMuraru peels contain flavonoids, phenols, and tannins that can help with inflammation, diabetes mellitus, and cancer. They can also lower cholesterol and bad cholesterol, and they can eliminate bacteria [<a href=\"#r-34\">34-35</a>], which makes them useful for herbal medicine and nutraceutical use. The most abundant compounds, terpenoids, and triterpenoids, in<em> M. sapientum</em> var. Muraru peels exert multifaceted effects on tumor development, inhibiting initiation, promoting apoptosis, and suppressing angiogenesis. These compounds also offer diverse biological activities, including antioxidant, anti-inflammatory, and cardiovascular protective effects [<a href=\"#r-36\">36-37</a>]. The peel's rich plant oils, containing lipids and therapeutic components like fatty acids and steroids, contribute to its potential to manage diseases. Secondary metabolites such as flavonoids, alkaloids, terpenoids, tannins, and phenols, found in abundance, greatly influenced the antioxidant activity of Muraru peels. Several studies [<a href=\"#r-38\">38-39</a>] have found these compounds to play a significant role as antioxidants. Oxidative stress deregulates cellular functions, leading to neurodegenerative diseases, gastro-duodenal pathogenesis, cancer, premature aging, inflammation, cardiovascular, and endocrine/metabolic dysfunction [<a href=\"#r-40\">40-41</a>]. Therefore, exogenous antioxidants like those found in Muraru peels can balance reactive oxygen species, preventing their occurrence and enhancing the management of chronic diseases and their complications. <br />\r\nIn our current study, the peels did not exhibit any signs of toxicity in acute toxicity tests, with an LD<sub>50</sub> greater than the limit dose of 2000 mg/kg. According to the classification by the Ministry of Agriculture, Food and Fisheries [<a href=\"#r-42\">42</a>], substances with an LD<sub>50</sub> above 2000 mg/kg are considered non-toxic. Furthermore, repeated doses of Muraru peel hydroethanolic extract administered over 28 days showed no signs of morbidity and no mortality. Furthermore, the rats' behavior patterns, skin, fur, eyes, salivation, and occurrence of diarrhea showed no discernible changes. Throughout the acute and subacute toxicity studies, the animals consistently demonstrated weight gain. Weight loss is recognized as a prominent signal for health issues and excessive toxicity [<a href=\"#r-43\">43</a>]. Importantly, the peel hydroethanolic extract induced no significant effects on the relative organ weights of the treated groups compared to the control group. The assessment of relative organ weights is crucial for identifying potential organ damage due to exposure to toxic substances. The extent of toxicity and the ratio of body weight would alter the weight of a damaged organ [<a href=\"#r-44\">44</a>]. These reflect the overall safety profile of banana peel hydroethanolic extract. <br />\r\nThe hemopoietin system functions as a sensitive target for toxic compounds and serves as a key indicator of physiological and pathological status in both humans and animals. During both acute and subacute tests, the banana peel hydroethanolic extract did not significantly alter the red blood cell (RBC) indices, suggesting that it has no effect on erythropoiesis, red blood cell morphology, or osmotic fragility. White blood cells (WBCs) play a crucial role in defending against infectious agents, tissue injury, or inflammation. Importantly, no significant changes were noted in neutrophils, lymphocytes, and monocytes in response to the banana peel extract, suggesting that the Muraru peel extract did not challenge the immune system of the animals in both acute and subacute tests. All of these results show that the hydroethanolic extract of the Muraru peel has a calming effect on hematological parameters, indicating that it may be safe for blood-related functions. Given that the analysis of hematological parameters provides valuable insights into blood-related functions and aids in diagnosing conditions such as anemia, infections, acute hemorrhagic states, allergies, and immune deficiencies [<a href=\"#r-45\">45</a>].<br />\r\nAssessing liver and kidney function is crucial to evaluating the potential toxicity of herbal formulations and drugs [<a href=\"#r-46\">46</a>]. Urea serves as an indicator of kidney function, while creatinine is employed as a marker for the glomerular filtration rate. ALT is a sensitive marker for detecting liver cell damage, and AST is present in RBCs, heart, kidney, and skeletal muscles [<a href=\"#r-46\">46</a>]. These parameters were examined to determine potential alterations induced by the peel hydroethanolic extract. The absence of significant changes in ALT, AST, creatinine, and urea levels, recognized indicators of liver and kidney functions, suggests that the acute and subacute administration of the extract did not adversely affect hepatocytes, kidneys, or the normal metabolism of the rats. The safety of the extract is further evident in the morphology of the organs, which displayed no alterations or lesions. <br />\r\nHence, the wealth of bioactive compounds found in <em>M. sapientum</em> var. Muraru peels, coupled with their high antioxidant capacity and safety profile, positions them as valuable candidates for nutraceutical applications. These versatile peels could potentially be utilized in various functional foods, dietary supplements, or natural health products aimed at promoting wellness and preventing oxidative stress-related diseases.</p>"
},
{
"section_number": 5,
"section_title": "CONCLUSION",
"body": "<p><em>M. sapientum</em> var Muraru unripe and ripe peels are rich in alkaloids, phenols, terpenoids, flavonoids, tannins, and phytosterols. The peels exhibit significant antioxidant activity, supported by an IC<sub>50</sub> indicating moderate to strong activity. The results of acute and subacute oral toxicity studies on <em>M. sapientum</em> var Muraru peel hydroethanolic extract indicate its non-toxic nature across all tested groups, with no observable morbidity. These findings suggest that the peels could serve as a valuable source of bioactive compounds and can be efficiently utilized, minimizing waste. These findings position Muraru peels as promising candidates for pharmacological and therapeutic applications as nutraceuticals in improving oxidative stress.</p>"
},
{
"section_number": 6,
"section_title": "ACKNOWLEDGEMENTS",
"body": "<p>The authors appreciate Mr. John Kumau (Gok-Botany Lab) for plant identification, the GoK Chemistry Laboratory at JKUAT for assisting in the phytochemical analysis, and the Analytical Laboratory for the GC-MS. Special thanks are extended to Patricia Karanja, the Muraru banana farmer, and Mr. Perminus Kimathi of the Small Animal Facility for Research and Innovation (SAFARI) at JKUAT for their assistance with animal handling. Heartfelt gratitude is extended to Dr. Ifeoluwa Gbala, who provided valuable insights, critical feedback, and dedicated time to help shape the content. A huge thank you to the African Union for funding the research project through the Pan African University scholarship.</p>"
},
{
"section_number": 7,
"section_title": "AUTHOR CONTRIBUTIONS",
"body": "<p>MCM: worked on methodology, data curation, conceptualization visualization, validation, writing the original draft, writing the review, and editing. AK: visualization, validation, methodology, review, and editing. RW: supervision, conceptualization, methodology, review, and editing. All authors approved the final version of the manuscript.</p>"
},
{
"section_number": 8,
"section_title": "CONFLICTS OF INTEREST",
"body": "<p>There is no conflict of interest among the authors.</p>"
}
],
"figures": [
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/58/09/280-1708590809-Figure1.jpg",
"caption": "Figure 1. GC-MS chromatogram of Muraru unripe and ripe peel extracts. A) Unripe ethanolic extracts showing the presence of undecane (alkane), hexadecenoic acid, methyl ester (saturated fatty acid), 9,12-octadecadienoic acid, methyl ester (unsaturated fatty acid), methyl 8,11,14-heptadecatrienoate (fatty acid), thunbergol (triterpenoid), and 9,19-Cycloergost-24(28)-en-3-ol, 4,14-dimethyl-, acetate, (3β,4α,5α)- (terpenoids). B) Unripe methanolic extracts showing the presence of 2,4-Di-tert-butylphenol (phenols), Pyrrolidine-2-carboxylic acid, methyl-phenyl-amide (saturated heterocycle), Decanedioic acid, bis(2-ethylhexyl) ester (fatty acid), 9,19-Cyclolanostan-3-ol, 24-methylene-, (3β)- (triterpenoid) and 9,19-Cycloergost-24(28)-en-3-ol, 4,14-dimethyl-, acetate, (3β,4α,5α)- (terpenoids). C) Ripe ethanolic extracts showing the presence of melezitose (trisaccharide), Hexadecanoic acid, ethyl ester (saturated fatty acid), 3,7,11,15-Tetramethyl-2-hexadecen-1-OL (phytol), 9,19-Cycloergost-24(28)-en-3-ol, 4,14-dimethyl-, acetate, (3β,4α,5α)- (terpenoids) and Androst-1-en-3-one, 4,4-dimethyl-, (5alpha)- (steroid). D) Ripe methanolic 14-methyl-pentadecanoic acid, methyl ester (fatty acid), 9,12-Octadecadienoic acid, methyl ester (unsaturated fatty acid), 9,12,15-Octadecatrienoic acid, methyl ester (lipid), Heptadecanoic acid, 15-methyl-, methyl ester (saturated fatty acid) and alpha. Tocopheryl acetate (vitamin E).",
"featured": false
},
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/58/09/280-1708590809-Figure2.jpg",
"caption": "Figure 2. The colorimetric DPPH antioxidant assay revealed that Muraru peels have significant antioxidant capacity. Total antioxidant capacity varied with the peel state, with the unripe peels in both solvents revealing the highest scavenging percentage compared to the ripe peel state. The scavenging percentage of the peel extracts against free radical DPPH was 70.12%, 63.99%, 59.91%, and 48.06% at the highest concentration (200 μg/ml) for unripe methanolic, unripe hydroethanolic, ripe peel methanolic, and ripe peel hydroethanolic extracts, respectively. Ascorbic acid was used as the standard for the assay, which also showed that the scavenging percentage increases with an increase in concentration, with 200 μg/mL being the optimum concentration.",
"featured": false
},
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/58/09/280-1708590809-Figure3.jpg",
"caption": "Figure 3. The effect of Muraru peel hydroethanolic extract on body and organ weights during the acute and subacute toxicity tests. A) The mean weight of animals during the acute toxicity study. B) The mean body weight of Wistar rats during the subacute study. C) The relative organ weight obtained after the acute toxicity study. D) The relative organ weight obtained after the subacute toxicity study. The organs in both acute and subacute toxicity tests during harvesting did not show any change in color or enlargement. Relative organ weight was calculated as a percentage of the absolute weight. Values are expressed as a mean ± SEM (standard error mean). The error bars represent SEM (n = 3 rats/group).",
"featured": false
},
{
"figure": "https://plant-trends.bsmiab.org/media/article_images/2024/58/09/280-1708590809-Figure4.jpg",
"caption": "Figure 4. Morphologies of various organs following sub-acute toxicity. Micrographs of lung, liver, heart, spleen and kidney after 28 days of administration of hydroethanolic Muraru peel extracts. Hematoxylin and eosin stain (Magnification 400×).",
"featured": true
}
],
"authors": [
{
"id": 16,
"affiliation": [
{
"affiliation": "Department of Molecular Biology and Biotechnology, Pan African University Institute for Basic Sciences Technology, and Innovation (PAUSTI), Juja, Kenya"
}
],
"first_name": "Mishell Chantelle",
"family_name": "Mahora",
"email": "michellemahora@gmail.com",
"author_order": 1,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": true,
"corresponding_author_info": "Mishell Chantelle Mahora\r\nDepartment of Molecular Biology and Biotechnology, Pan African University Institute for Basic Sciences Technology, and Innovation (PAUSTI), Juja, Kenya; Email: michellemahora@gmail.com",
"article": 6
},
{
"id": 17,
"affiliation": [
{
"affiliation": "Department of Pharmacy, Jomo Kenyatta University of Agriculture and Technology (JKUAT), Juja, Kenya"
}
],
"first_name": "Alex",
"family_name": "Kigundu",
"email": null,
"author_order": 2,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 6
},
{
"id": 18,
"affiliation": [
{
"affiliation": "Department of Zoology, JKUAT, Juja, Kenya"
}
],
"first_name": "Rebecca",
"family_name": "Waihenya",
"email": null,
"author_order": 3,
"ORCID": null,
"co_first_author": false,
"co_author": false,
"corresponding": false,
"corresponding_author_info": "",
"article": 6
}
],
"views": 2597,
"downloads": 236,
"references": [
{
"id": 167,
"serial_number": 1,
"reference": "Bhavani M, Morya S, et al. Nutraceutical applications of banana peel, International Journal of Food Properties. 2023; 26(1):1277–1289.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 168,
"serial_number": 2,
"reference": "Jenkins W, Tucker ME, et al. Routledge handbook of religion and ecology, Routledge Handbook of Religion and Ecology. 2016.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 169,
"serial_number": 3,
"reference": "Onyango M, Karamura D, et al. Morphological Characterisation of East African AAB and AA Dessert Bananas (Musa spp), Proc. Int’l ISHS-ProMusa Symp. 2011.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 170,
"serial_number": 4,
"reference": "Ishak NA, Yusuf M, et al. Antidiabetic and Antioxidant Capacities of Local Banana Peels Extract by Using Subcritical Water Extraction Technique, The Malaysian Journal of Islamic. 2019; 26.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 171,
"serial_number": 5,
"reference": "Singh B, Singh JP, et al. Bioactive compounds in banana and their associated health benefits: A review, Food Chemistry. 2016; (16): 1-11.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 172,
"serial_number": 6,
"reference": "Avcil M. Role of Antioxidants and its Functions in Human Body, Oxidants and Antioxidants in Medical Science. 2022; 11(7): 1-2.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 173,
"serial_number": 7,
"reference": "Ohiagu OF, Chikezie P, et al. Toxicological Significance of Bioactive Compounds of Plant Origin, Pharmacogn. Commn. 2021;11(2):67-77.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 174,
"serial_number": 8,
"reference": "Guaadaoui A, Benaicha N, et al. What is Bioactive Compound? A combined definition for a preliminary consensus. International Journal of Nutrition and Food Sciences. 2014; 3(3): 174-179.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 175,
"serial_number": 9,
"reference": "Bernhoft A. A brief review on bioactive compounds in plants, Bioactive compounds in plants- benefits and risks for man and animals. 2010: 11–17.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 176,
"serial_number": 10,
"reference": "Mondal A, Banerjee, S. et al. Cancer Preventive and Therapeutic Potential of Banana and Its Bioactive Constituents: A Systematic, Comprehensive, and Mechanistic Review, Frontiers in Oncology. 2021; 11: 1-19.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 177,
"serial_number": 11,
"reference": "Soltani Firouz M, Alimardani R, et al. Evaluating banana ripening status from measuring dielectric properties, Journal of Food Engineering. 2011; 105: 625–631.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 178,
"serial_number": 12,
"reference": "Toupal S, Coşansu S. Antioxidant and antimicrobial properties of freeze-dried banana and watermelon peel powders, Food and Humanity. 2023; (1)607–613.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 179,
"serial_number": 13,
"reference": "Oyeyinka BO, Afolayan AJ. Potentials of Musa Species Fruits against Oxidative Factors and Dietary Secondary. 2020; 25, 5036.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 180,
"serial_number": 14,
"reference": "Siddique S, Nawaz S, et al. Phytochemical screening and in-vitro evaluation of pharmacological activities of peels of Musa sapientum and Carica papaya fruit, Natural Product Research, 2018; 6419.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 181,
"serial_number": 15,
"reference": "Kibria AA, Kar A. Extraction and Evaluation of Phytochemicals from Banana Peels (Musa Sapientum) And Banana Plants (Musa paradisiaca), Malaysian Journal of Halal Research Journal. 2019; 2(1): 22–26.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 182,
"serial_number": 16,
"reference": "Aboul-enein AM, Salama ZA. et al. Research Article Identification of phenolic compounds from banana peel (Musa paradaisica L.) as antioxidant and antimicrobial agents. 2016; 8(4): 46–55.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 183,
"serial_number": 17,
"reference": "Santhosh R, Suriyanarayanan B. Plants: A Source for New Antimycobacterial Drugs, Planta medica. 2013; 80.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 184,
"serial_number": 18,
"reference": "Chaudhary P, Varshney N. et al. Analysis of pharmacognostical standardization, antioxidant capacity and separation of phytocompounds from five different vegetable peels using different solvents, Environment Conservation Journal. 2022; 23: 247–259.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 185,
"serial_number": 19,
"reference": "Council N. R. National Research Council (US) Committee for the Update of the Guide for the Care and Use of Laboratory Animals. Guide for the Care and Use of Laboratory Animals, 8th edition Washington (DC). NAP: 2011.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 186,
"serial_number": 20,
"reference": "Organisation for Economic Co-operation and Development (OECD). Chapter IV, Guidelines for Toxicity Tests, 2008; 5–7.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 187,
"serial_number": 21,
"reference": "Navghare VV, Dhawale SC. Alexandria University Faculty of Medicine In vitro antioxidant, hypoglycemic and oral glucose tolerance test of banana peels, Alexandria Journal of Medicine, 2017; 53(3): 237–243.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 188,
"serial_number": 22,
"reference": "Pariyani R, Ismali IS. et al. Phytochemical Screening and Acute Oral Toxicity Study of Java Tea Leaf Extracts, BioMed Research International. 2015; 742420.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 189,
"serial_number": 23,
"reference": "Erhabor O, Muhammad H, et al. Interpretation of Full Blood Count Parameters in Health and Disease. 2021; 5(1): 00180.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 190,
"serial_number": 24,
"reference": "Dosseh K, Tchamdja E, et al. Anti-Fatigue Fatigue and Ergogenic Effect of Rourea Coccinea Schum. And Thonn. (Connaraceae) Ethanolic Extract in Rats. 2022; 14(11): 22759-22763.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 191,
"serial_number": 25,
"reference": "Shamaki B, Sandabe UK, et al. Toxicity studies and body weights changes in Wistar rats following oral administration of methanol extract from indigenous ganoderma sp. in Nigeria. 2017; 1(130): 138–141.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 192,
"serial_number": 26,
"reference": "Kamsu GT, Chuisseu D. et al. Toxicological Profile of the Aqueous Extract of Tectona grandis L. F. (Verbenaceae) Leaves: A Medicinal Plant Used in the Treatment of Typhoid Fever in Traditional Cameroonian Medicine, Journal of Toxicology. 2021; Article ID 6646771.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 193,
"serial_number": 27,
"reference": "Agung SFK, Febrianti M, et al. Comparison of Unripe Banana Peel of Kepok (Musa paradisiaca L.) and Klutuk (Musa balbisiana Colla): Phytochemical and Anti- dysenteriae Activity. 2020; 10(4): 911-914.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 194,
"serial_number": 28,
"reference": "González-Montelongo R, Lobo G, et al. Antioxidant activity in banana peel extracts: Testing extraction conditions and related bioactive compounds, Food Chemistry. 2010; 119: 1030–1039.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 195,
"serial_number": 29,
"reference": "Sundaram S, Anjum S. Antioxidant Activity and Protective effect of Banana Peel against Oxidative Haemolysis of Human Erythrocyte at Different Stages of Ripening, 2011; 1192–1206.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 196,
"serial_number": 30,
"reference": "Ehiowemwenguan G, Emoghene A, et al. Antibacterial and phytochemical analysis of Banana fruit peel. IOSR Journal of Pharmacy (IOSRPHR), 4, 18–25. https://doi.org/10.9790/3013-0408018025",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 197,
"serial_number": 31,
"reference": "Mordi R, Fadiaro T, et al. Identification by GC-MS of the Components of Oils of Banana Peels Extract, Phytochemical and Antimicrobial Analyses, Research Journal of Phytochemistry, 2016; (10):39–44.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 198,
"serial_number": 32,
"reference": "Puraikalan, Y, Current Research in Nutrition and Food Science Characterization of Proximate, Phytochemical and Antioxidant Analysis of Banana (Musa sapientum) Peels / Skins and Objective Evaluation of Ready to Eat / Cook Product made with Banana Peels, 2018; 06(2):362-391.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 199,
"serial_number": 33,
"reference": "Fidrianny I, Rizki R, et al. In vitro antioxidant activities from various extracts of banana peels using abts, Dpph assays and correlation with phenolic, Flavonoid, Carotenoid content, International Journal of Pharmacy and Pharmaceutical Sciences. 2014; 6: 299–303.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 200,
"serial_number": 34,
"reference": "Oriakhi K, Uadia P, Hypolipidemic Activity of Tetracarpidium conophorum (African walnut) Seed Oil and Its Mechanism of Action, Planta Med Int Open 2020; 7: 170–178.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 201,
"serial_number": 35,
"reference": "Harcourt P, Eedee F, et al. Antimicrobial Assessment of Fresh Ripe and Dry Ripe Musa sapientum L. Peels against Selected Isolates Associated with Urinary Tract Infection in, Port Harcourt. Journal of Medical Science and Clinical Research. 2018; 6(12): 420-430.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 202,
"serial_number": 36,
"reference": "Masyita A, Sari RM, et al. Terpenes and terpenoids as main bioactive compounds of essential oils, their roles in human health and potential application as natural food preservatives. Food Chemistry: X, 2022; (13): 100217.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 203,
"serial_number": 37,
"reference": "Yang W, Chen X, et al. Advances in Pharmacological Activities of Terpenoids, Natural Product Communications. 2020; 15(3): 1–13.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 204,
"serial_number": 38,
"reference": "Hikal WM Said-AL A, et al. Banana Peels: A Waste Treasure for Human Being. Evidence-Based Complementary and Alternative Medicine. 2022; Article ID 7616452.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 205,
"serial_number": 39,
"reference": "Madhavan M, Joy S. Antioxidant Activity of Fruit Peel Extracts of Musa paradisiaca L Antioxidant Activity of Fruit Peel Extracts of Musa paradisiaca L, Biological Forum International Journal. 2023; 15 (5): 278-282.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 206,
"serial_number": 40,
"reference": "Bennett V, Saliu IO. Exploring the Biochemical Components of Fresh Peels of Three Varieties of Musa sapientum (Banana). 2022; 10(4): 115-119.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 207,
"serial_number": 41,
"reference": "Sharifi-rad M, Kumar N. et al. Lifestyle, Oxidative Stress and Antioxidants: Back and Forth in the Pathophysiology of Chronic Diseases. Front. Physiol. 2020; 11:(694): 1–21.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 208,
"serial_number": 42,
"reference": "Ministry of Agriculture, Food and Fisheries, Pesticide Toxicity and Hazard, 2022; 1–9.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 209,
"serial_number": 43,
"reference": "Berlo V, Wouterson M. et al. 10 % Body weight (gain) change as criterion for the maximum tolerated dose: A critical analysis. 2021; 134: 105235.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 210,
"serial_number": 44,
"reference": "Ghauri AO, Mohiuddin E, et al. Acute and subacute toxicity studies of a poly herbal formulation used for diabetes. 2022; 38(6), 1668–1673.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 211,
"serial_number": 45,
"reference": "Celkan TT. What does a hemogram say to us? Turk pediatri arsivi. 2020; 55(2): 103–116.",
"pmc": null,
"DOI": null,
"article": 6
},
{
"id": 212,
"serial_number": 46,
"reference": "Ghauri AO, Mohiuddin E, et al. Acute and subacute toxicity studies of a poly herbal formulation used for diabetes., Pakistan journal of medical sciences, 2022; 38(6): 1668–1673.",
"pmc": null,
"DOI": null,
"article": 6
}
]
}
]
}
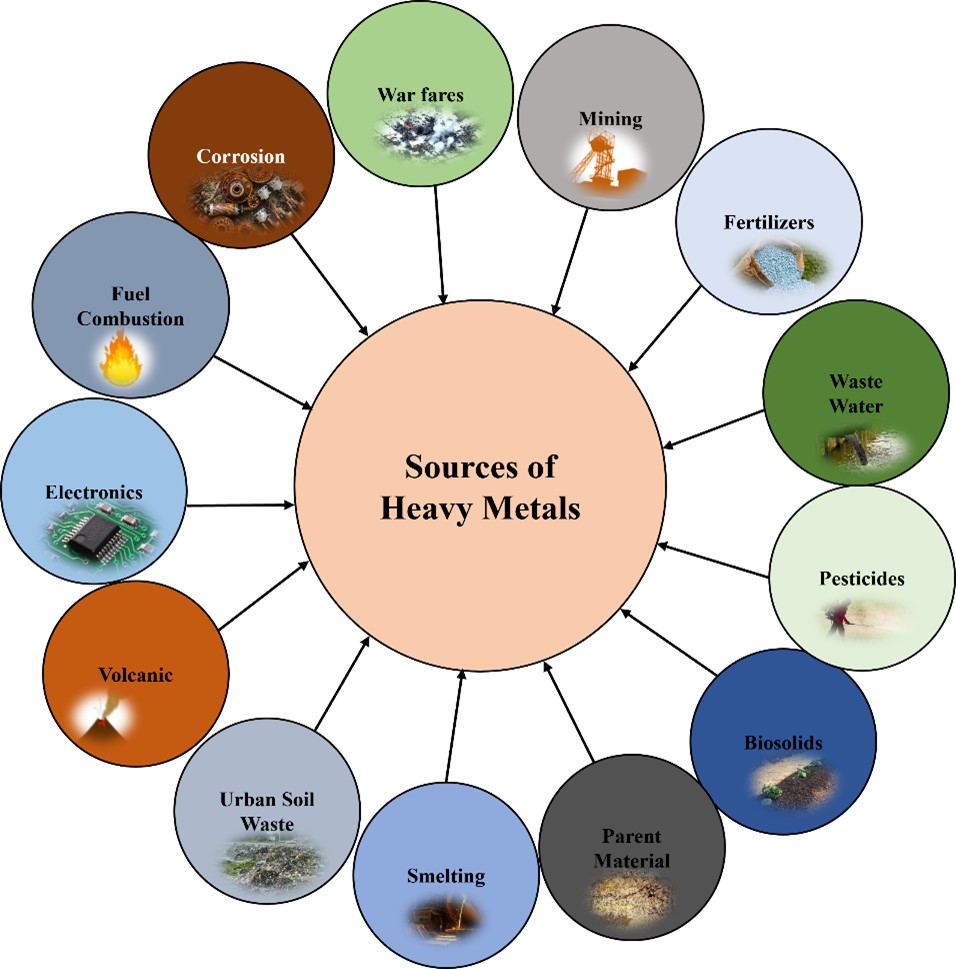{kind=link}
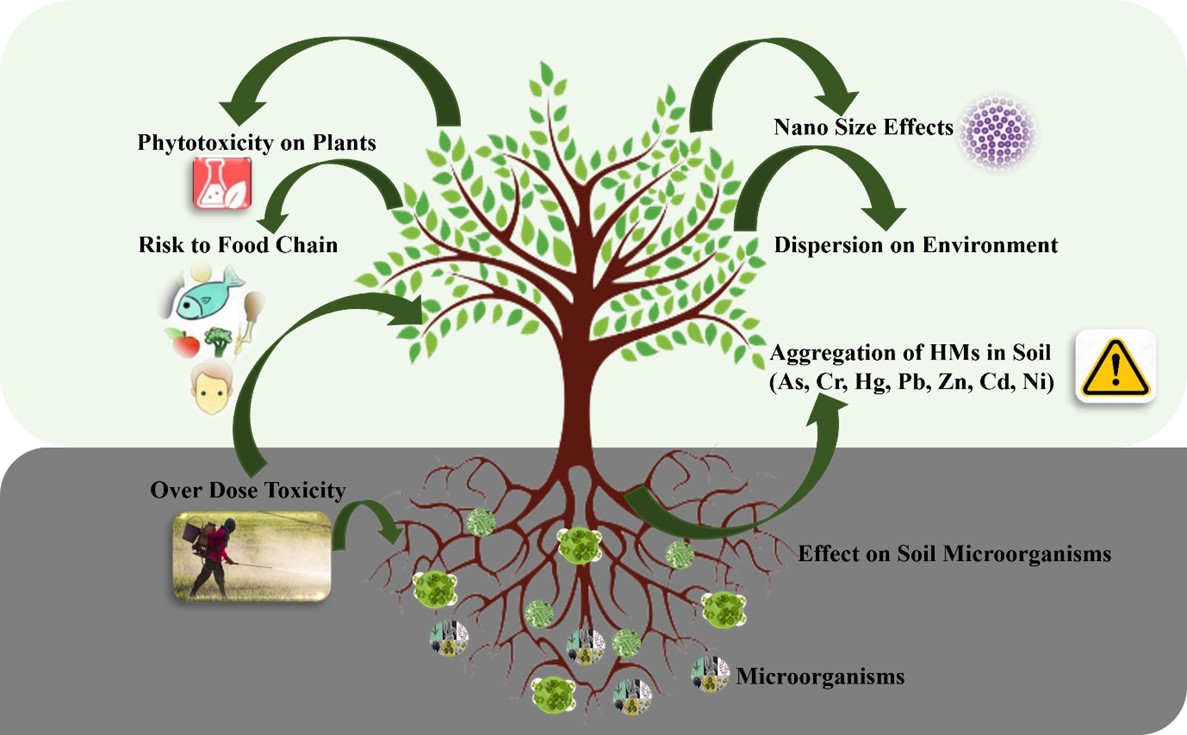{kind=link}
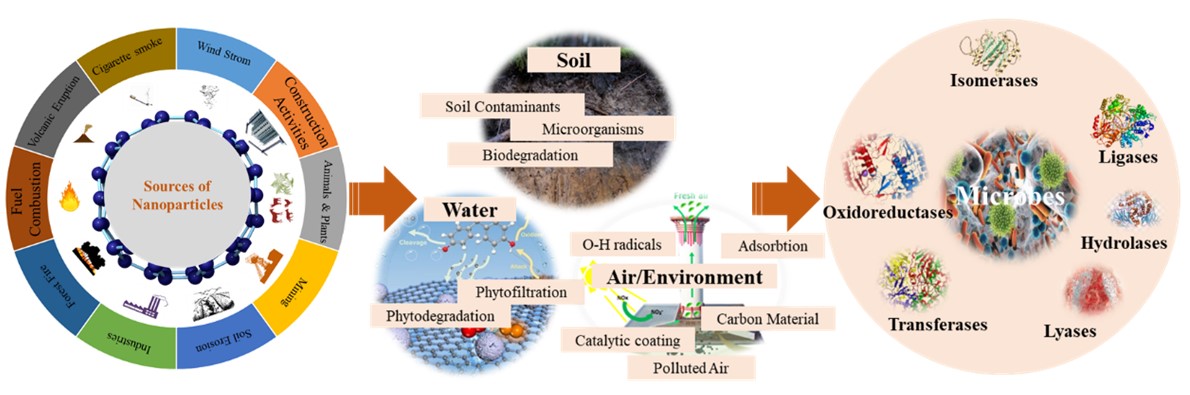{kind=link}
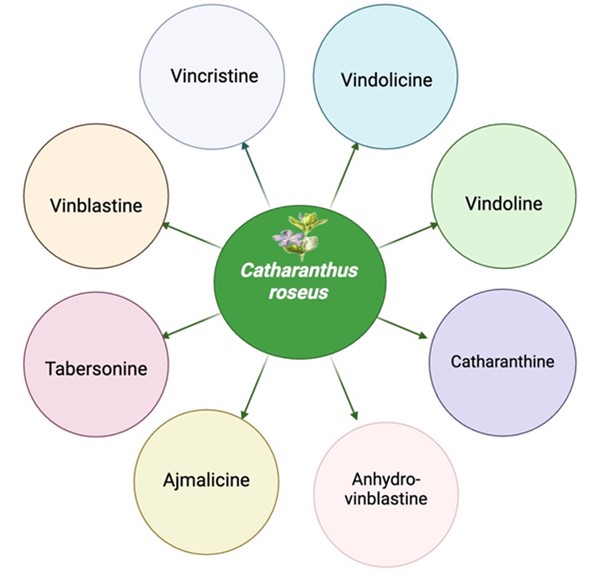{kind=link}
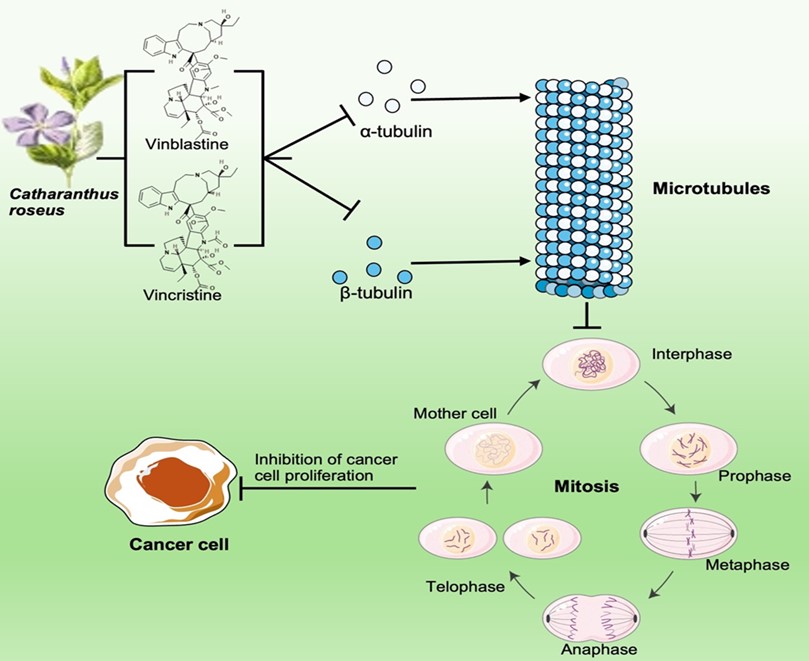{kind=link}
{kind=link}
{kind=link}
{kind=link}
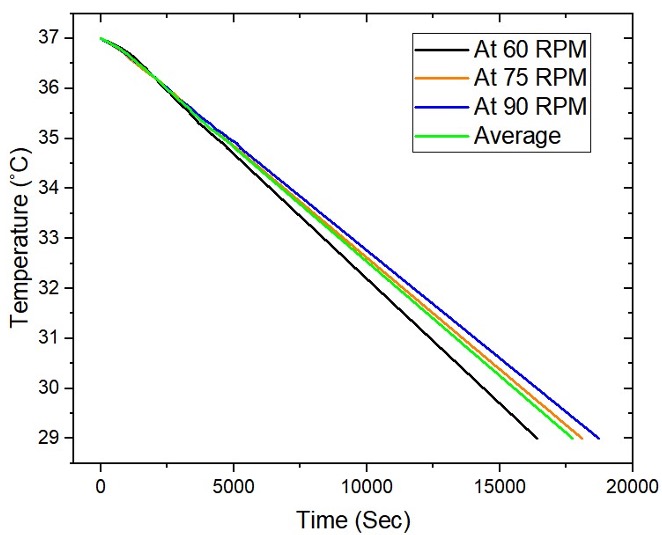{kind=link}
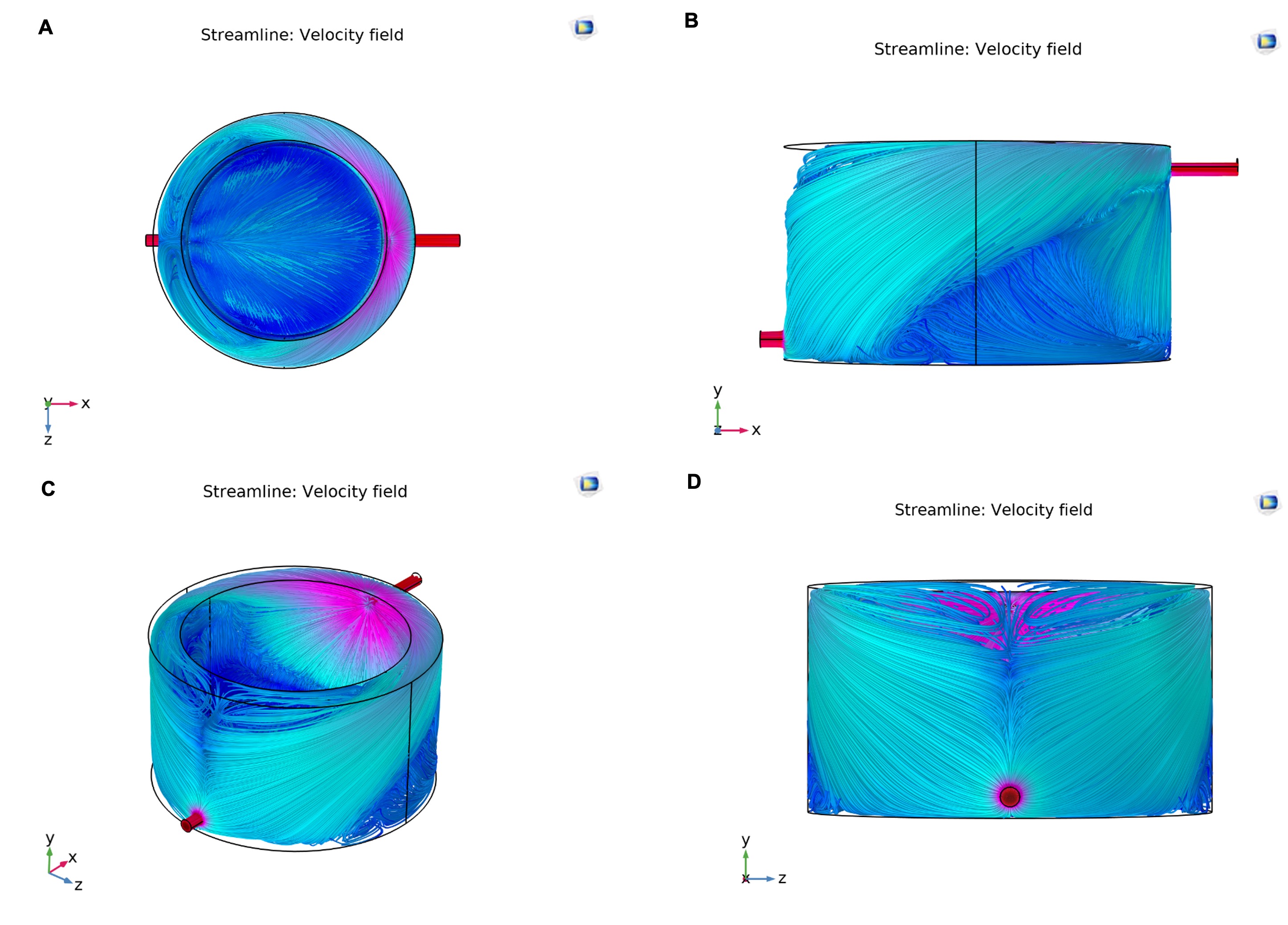{kind=link}
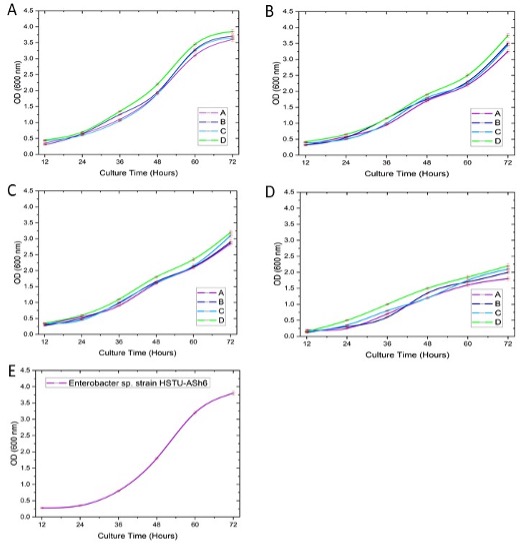{kind=link}
{kind=link}
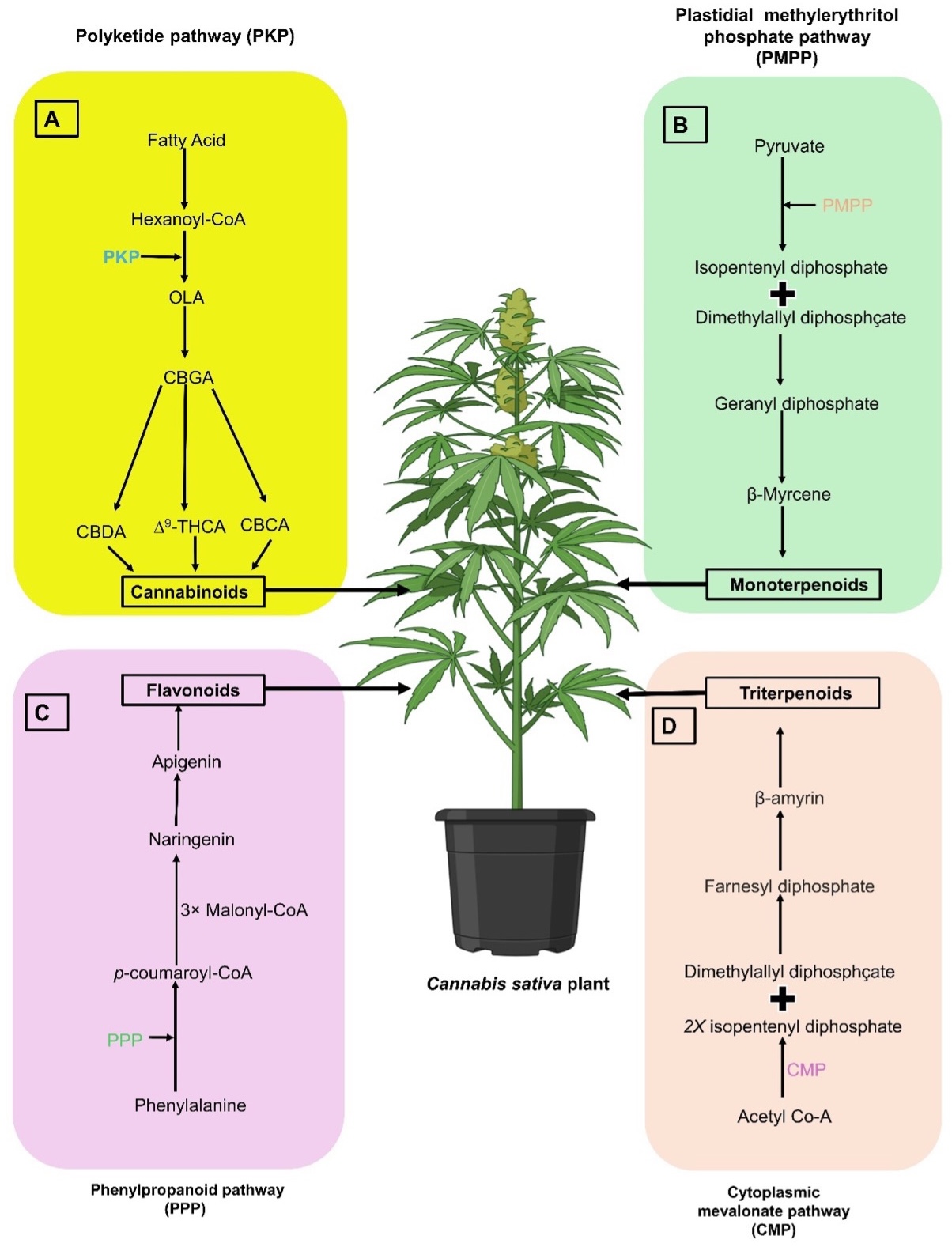{kind=link}
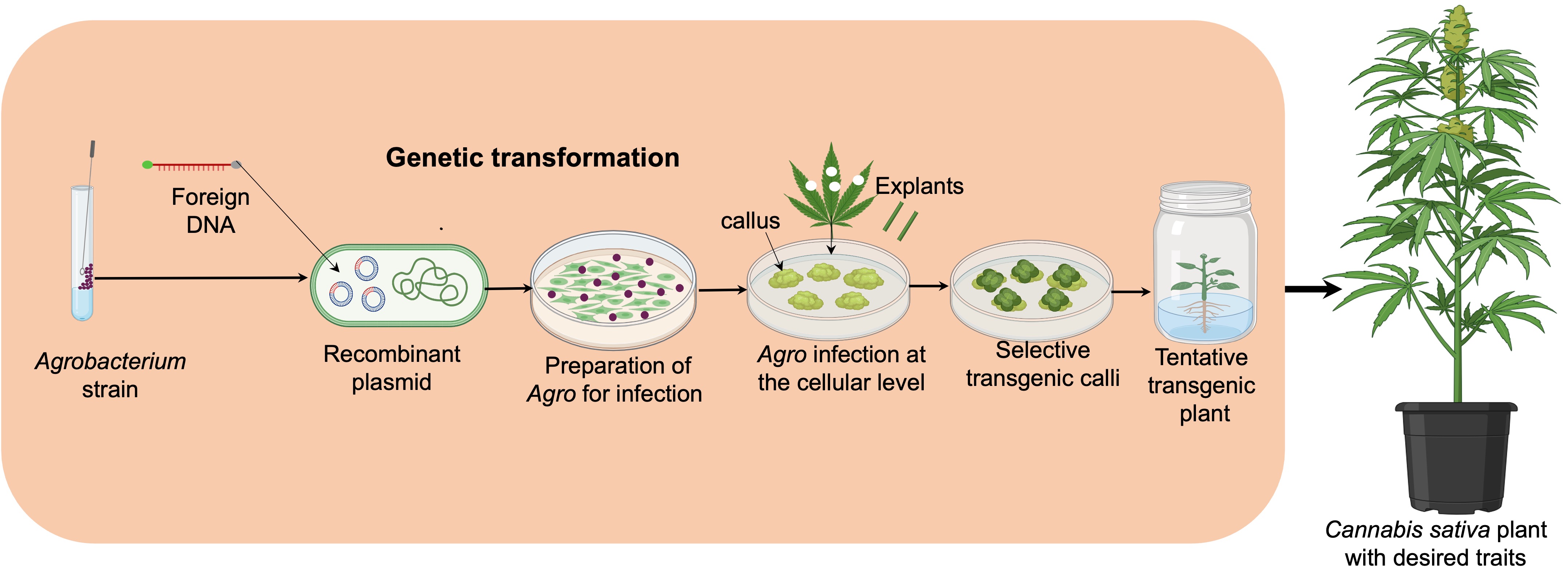{kind=link}
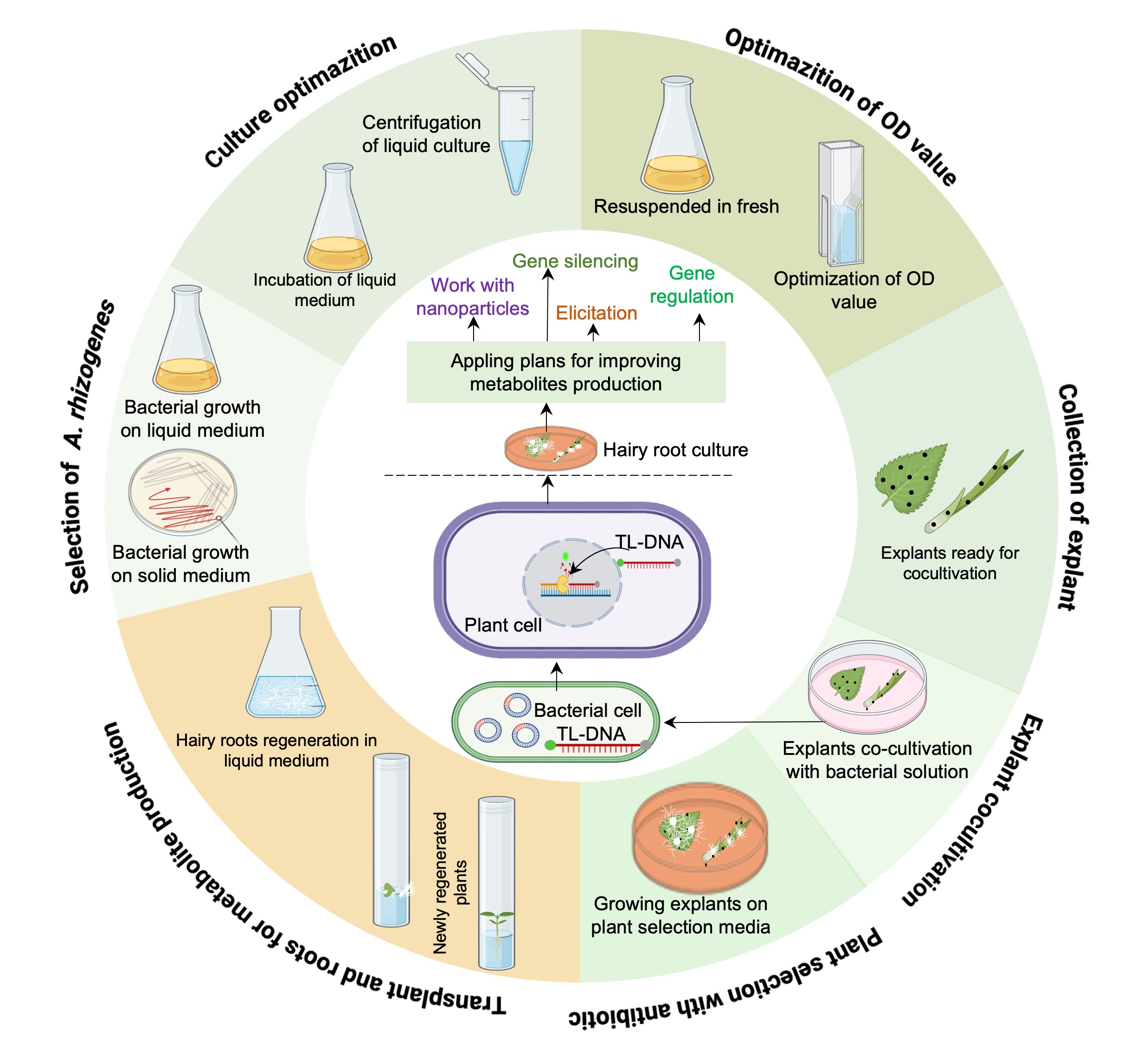{kind=link}
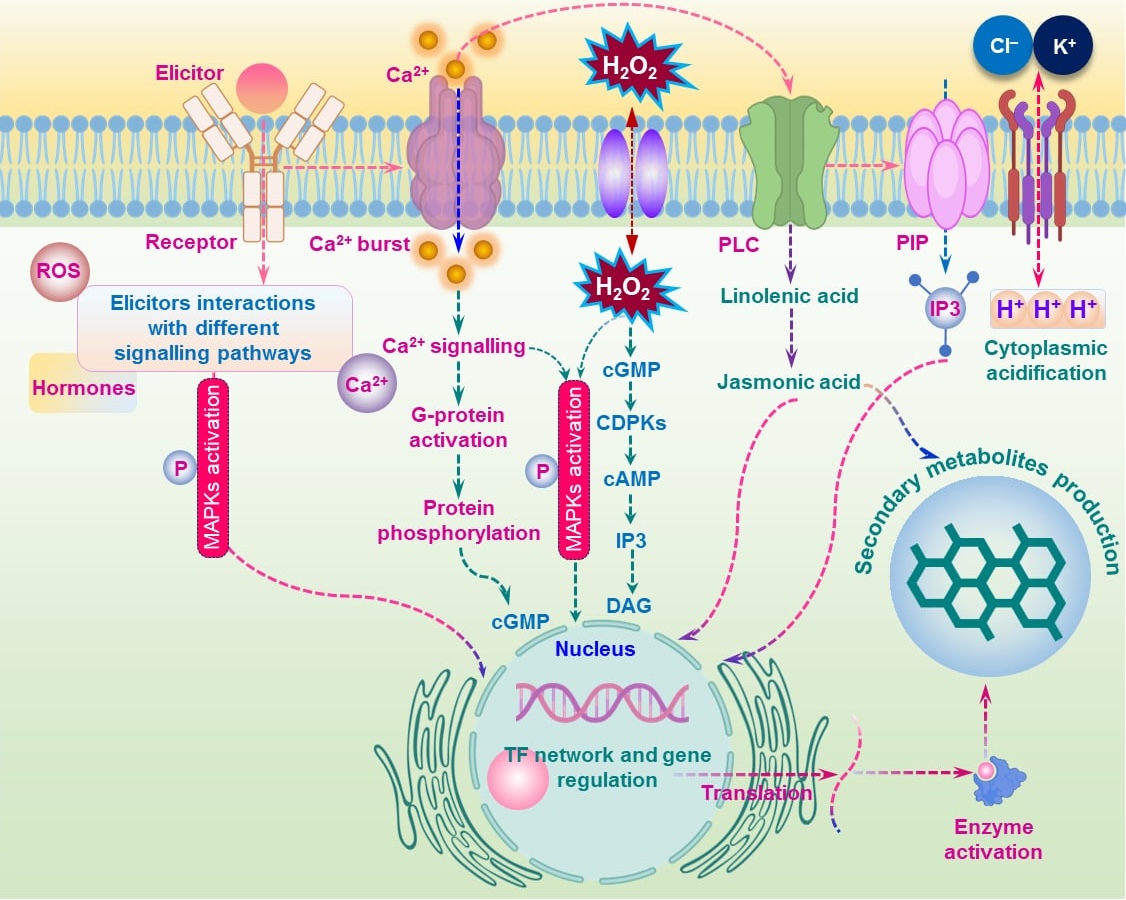{kind=link}
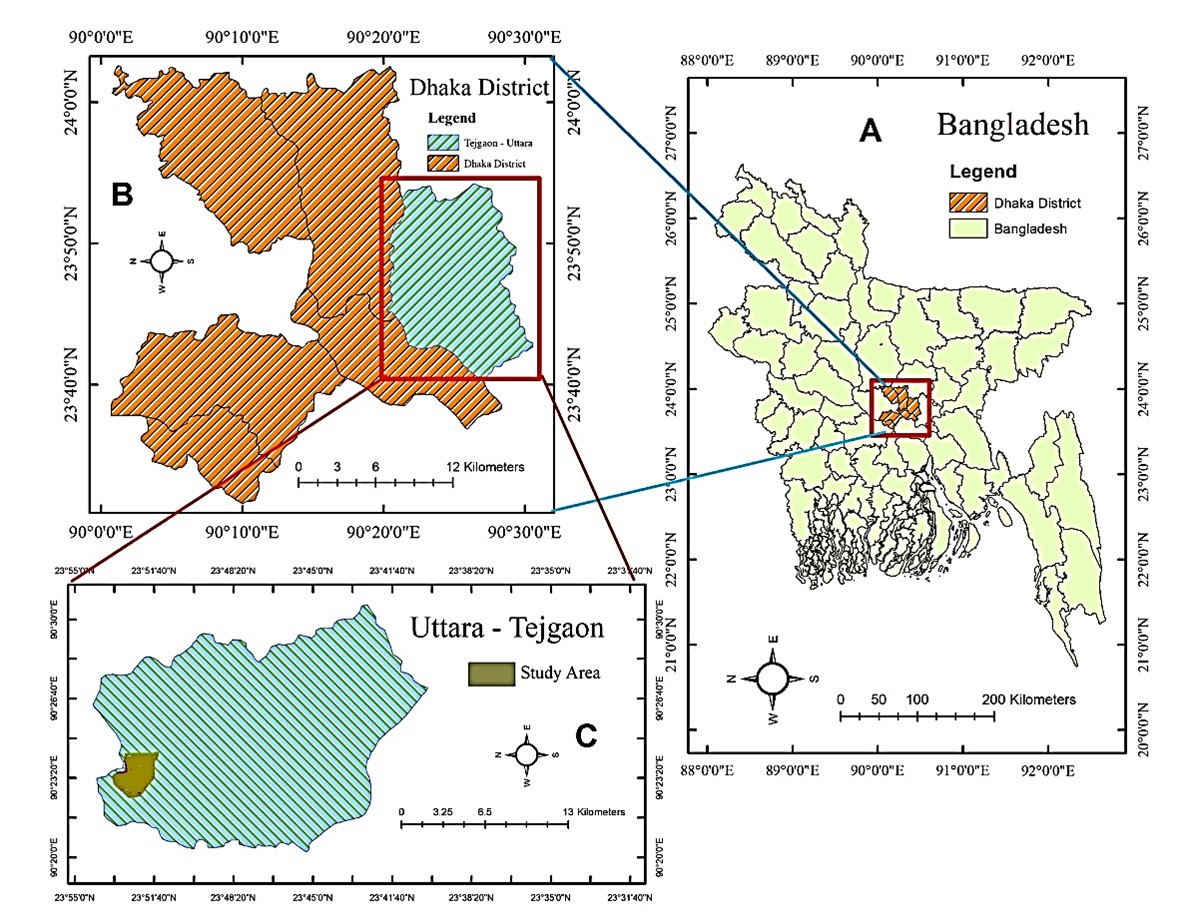{kind=link}
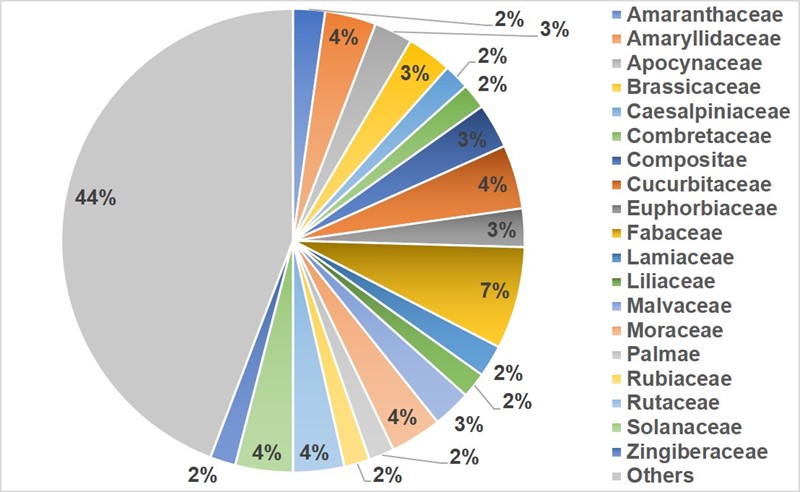{kind=link}
{kind=link}
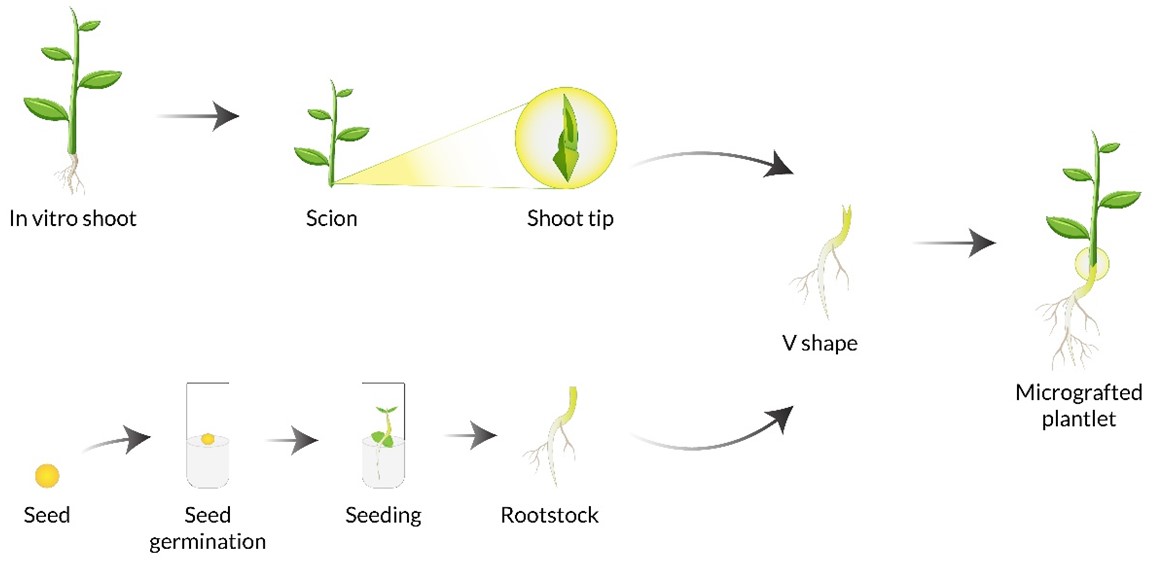{kind=link}
{kind=link}
{kind=link}
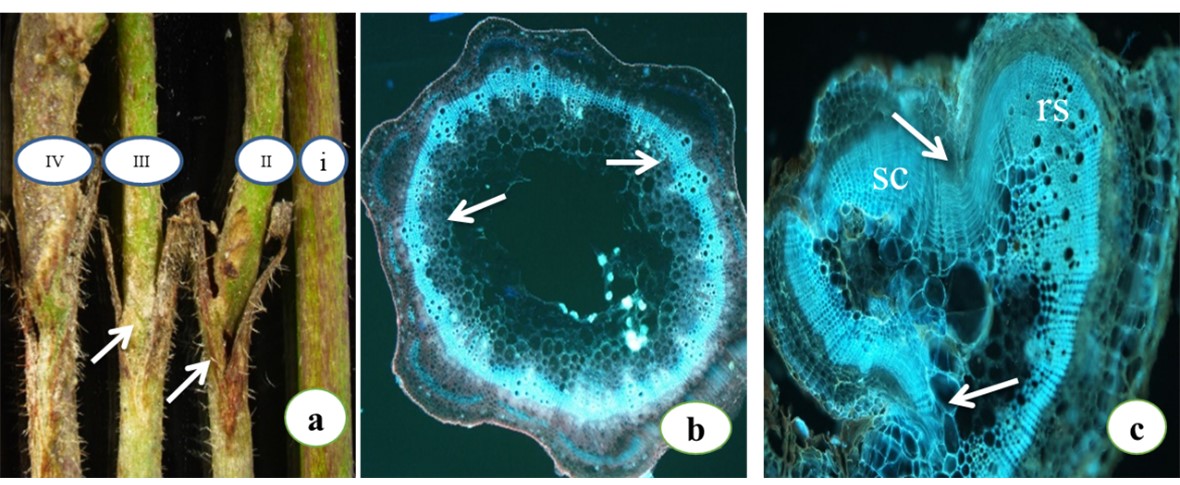{kind=link}
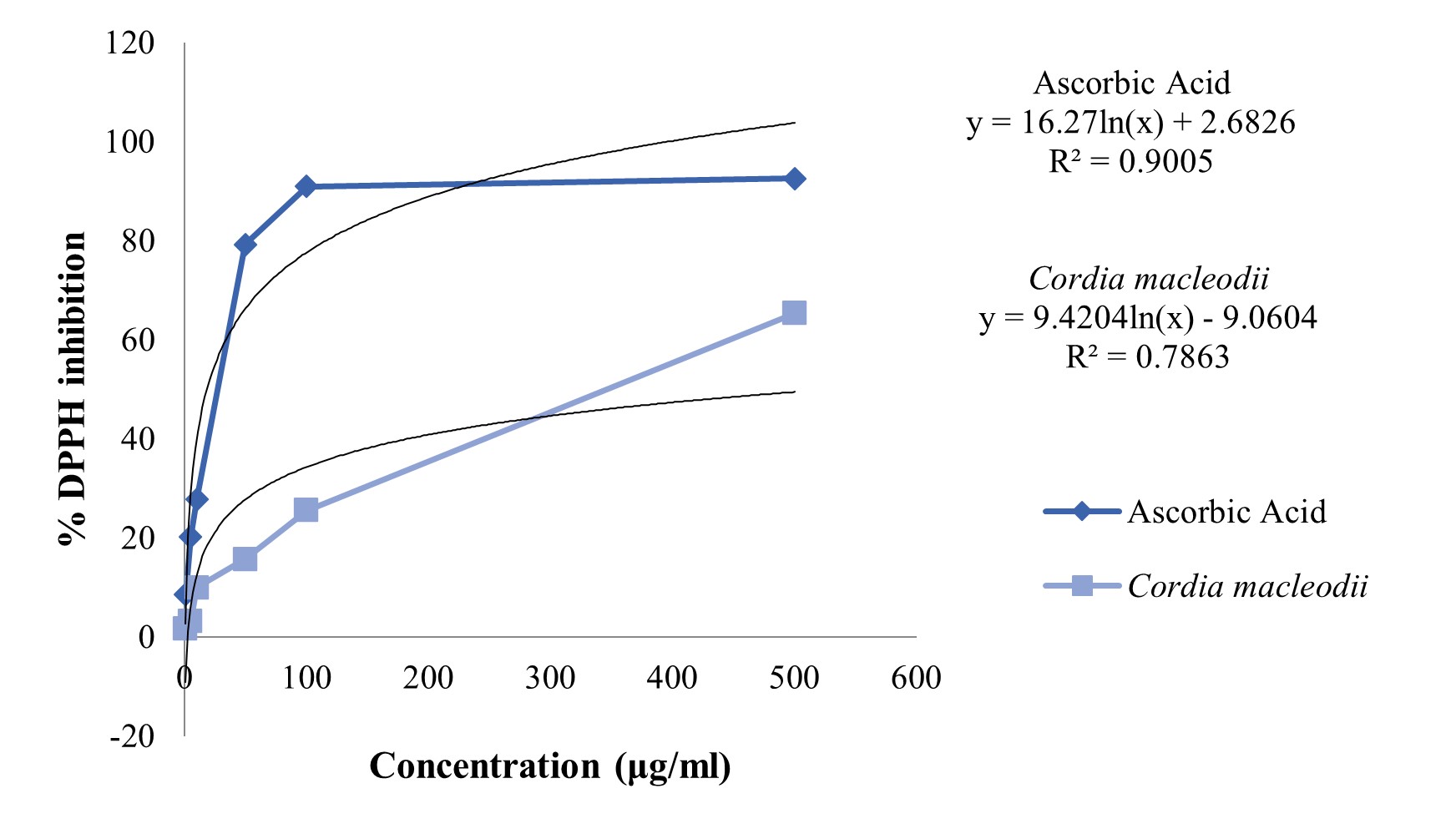{kind=link}
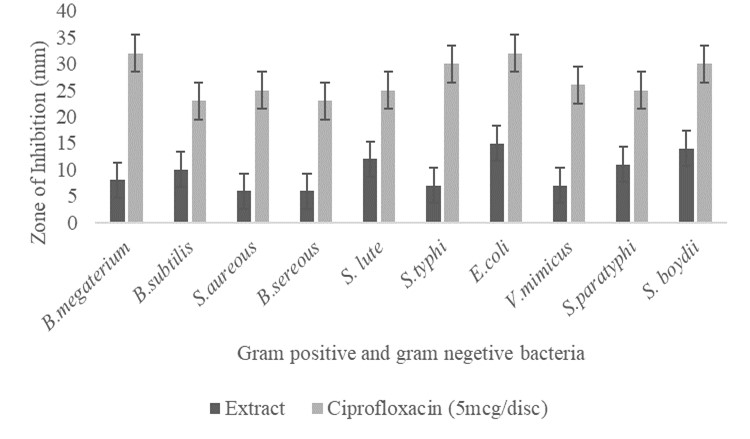{kind=link}
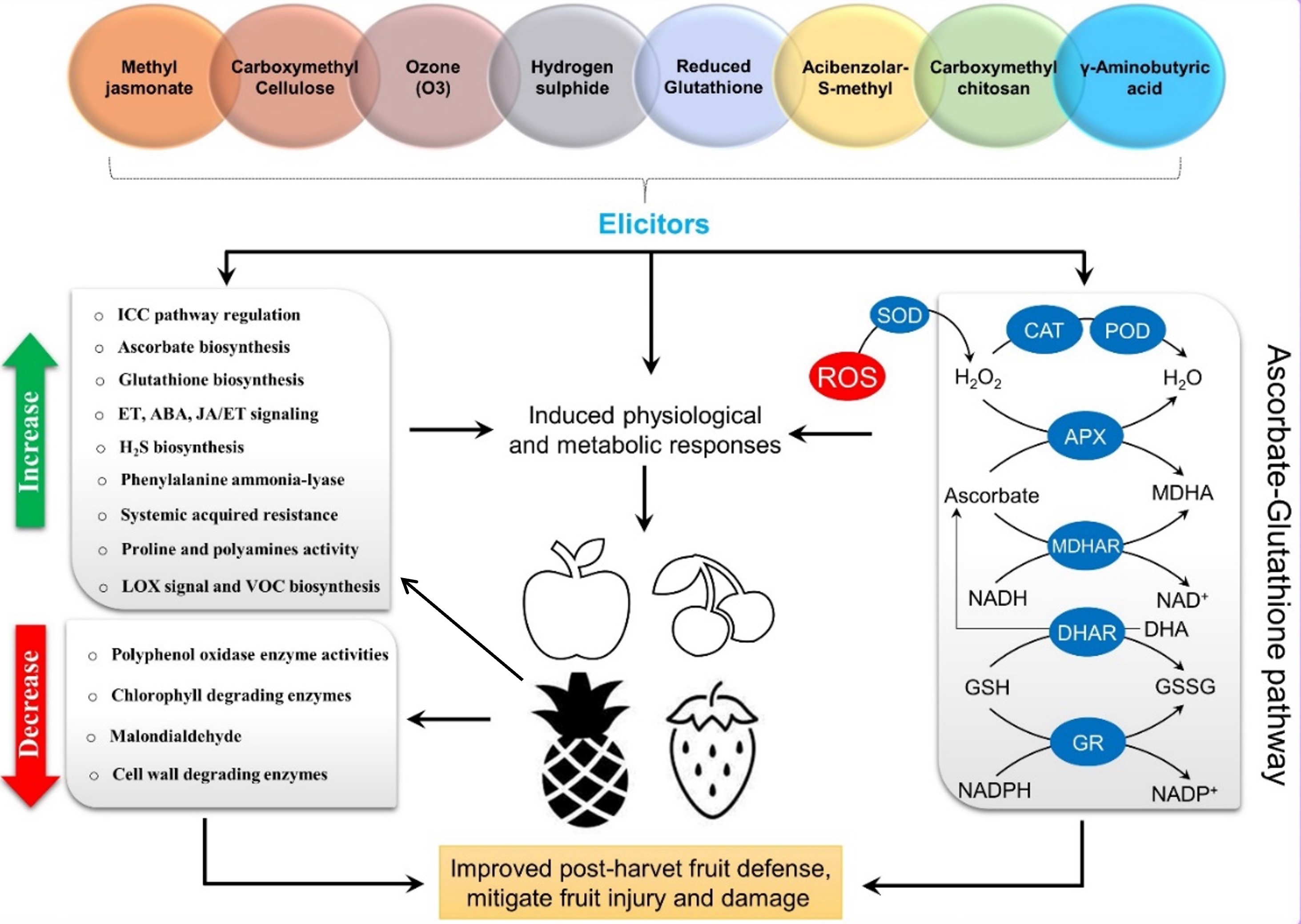{kind=link}
{kind=link}
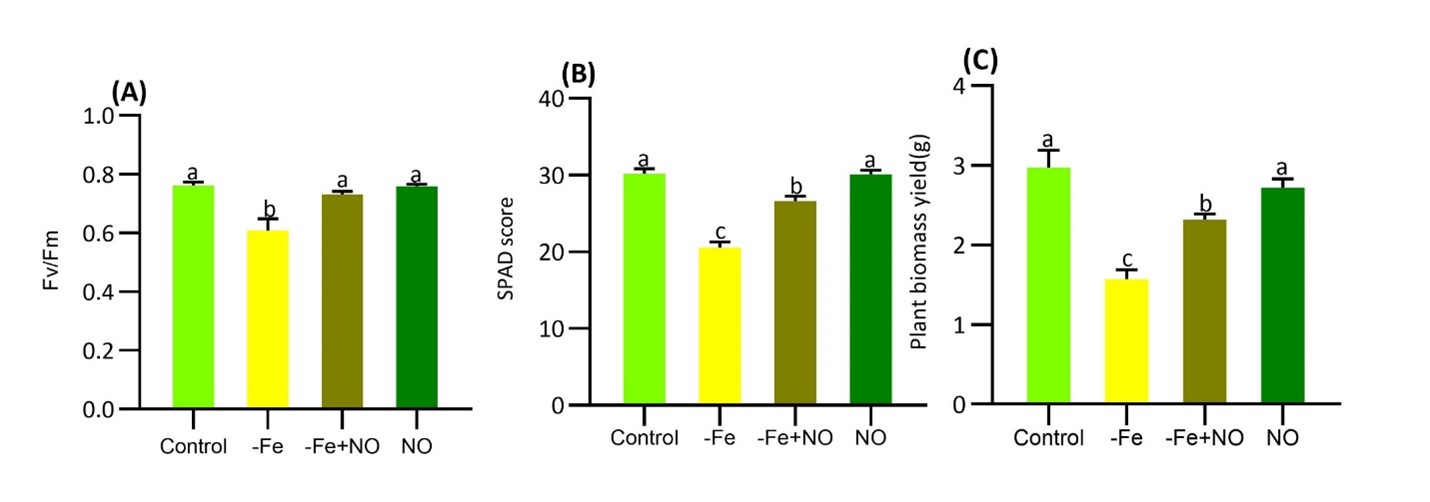{kind=link}
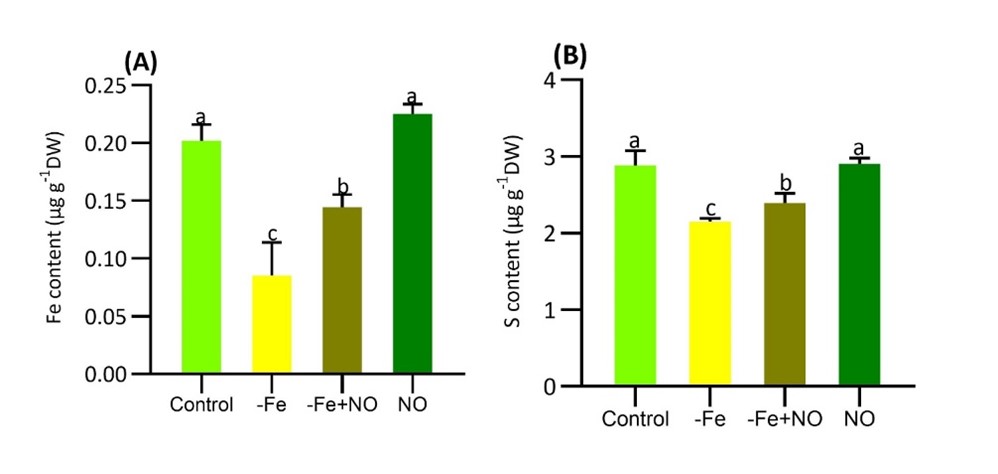{kind=link}
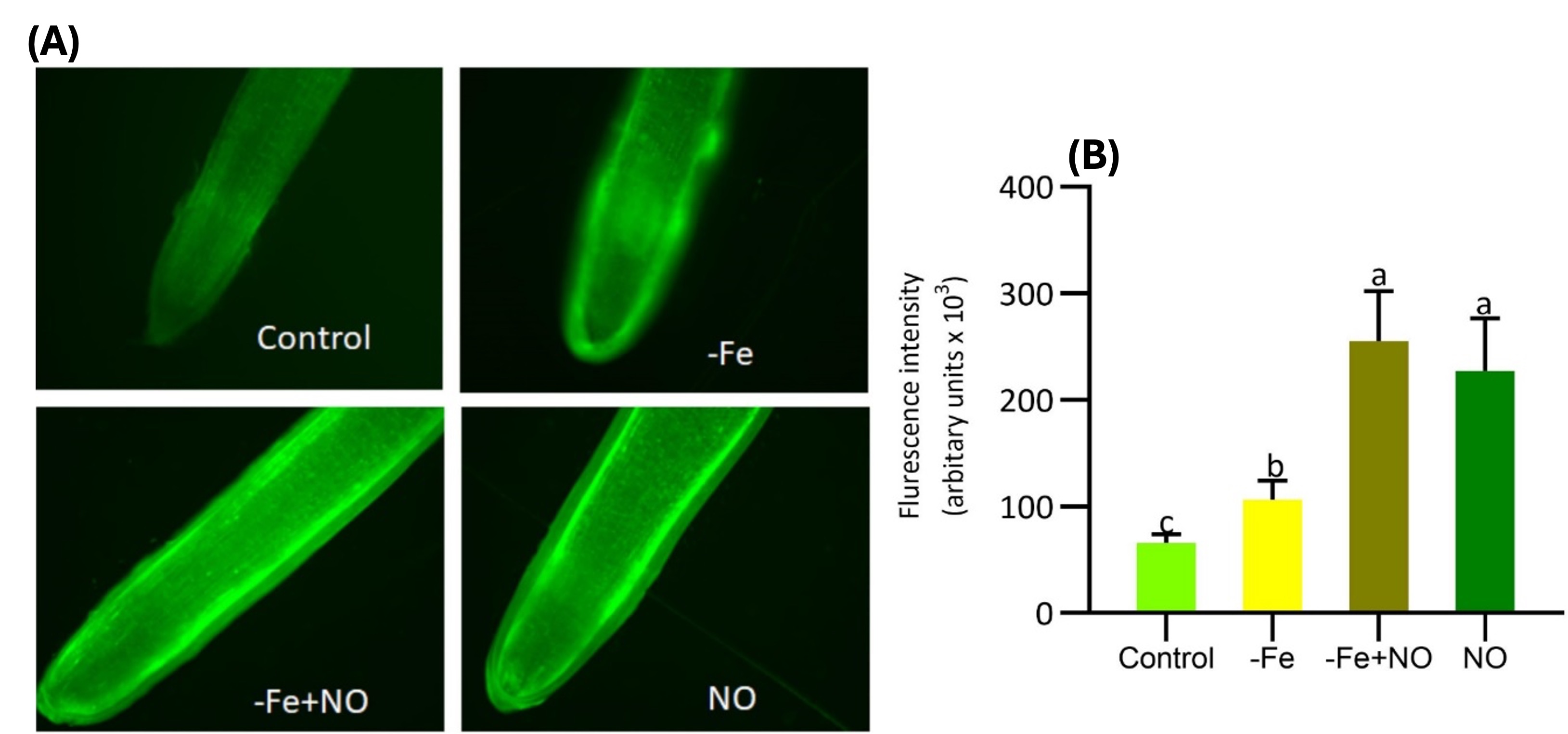{kind=link}
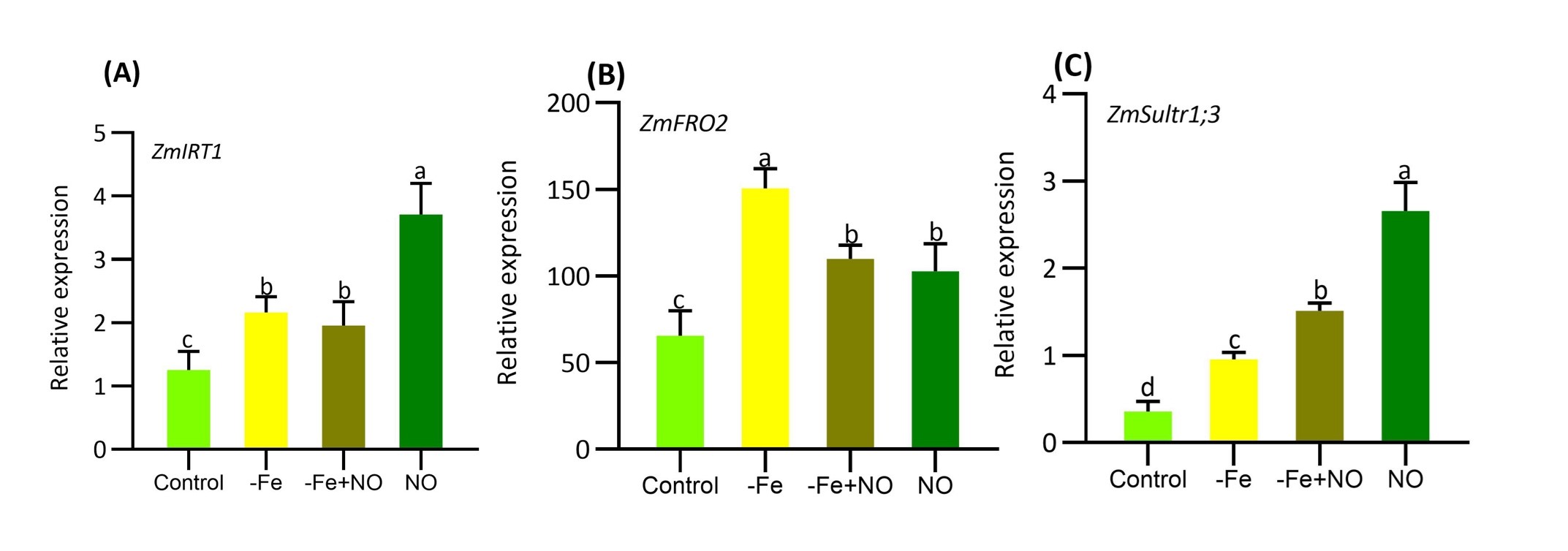{kind=link}
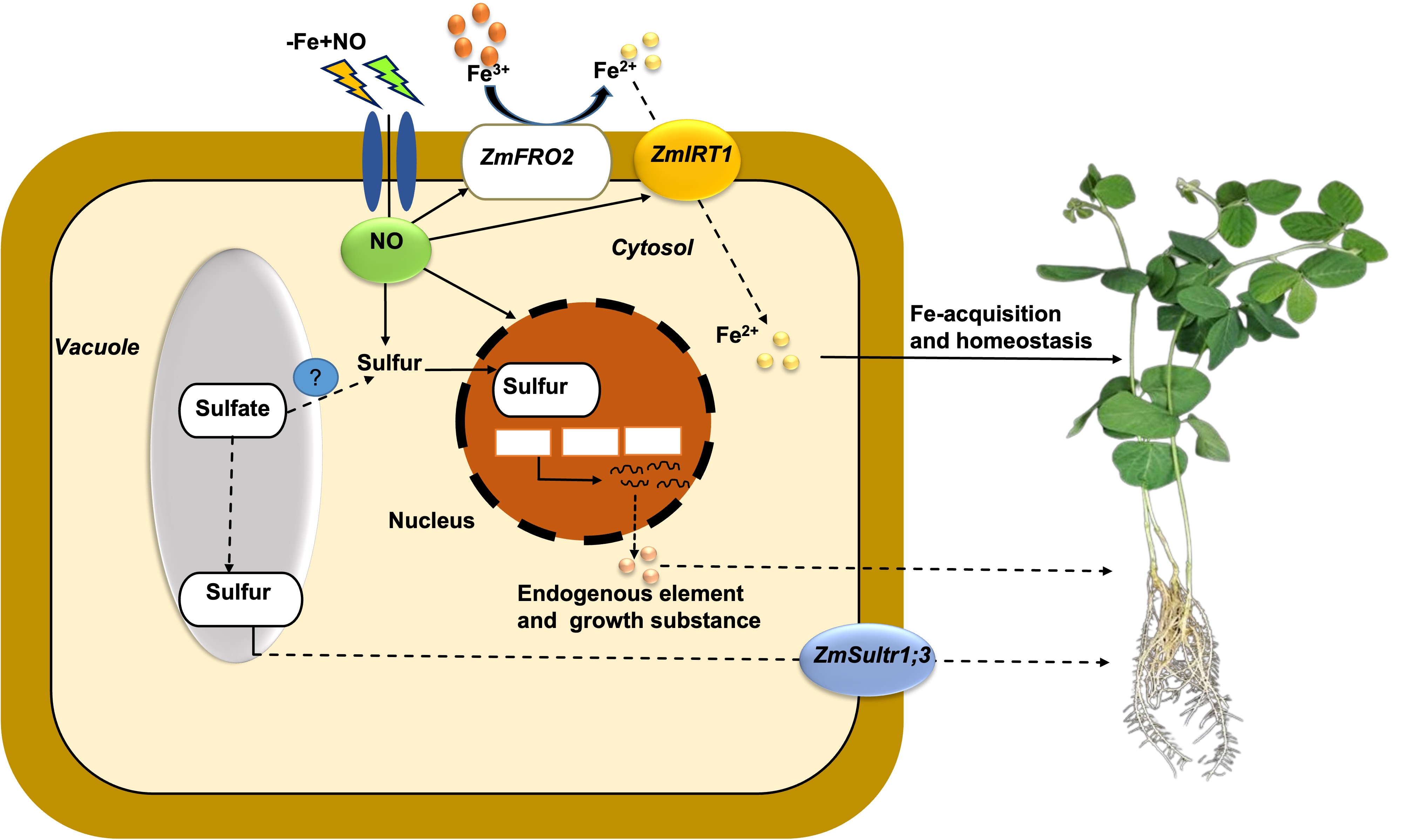{kind=link}
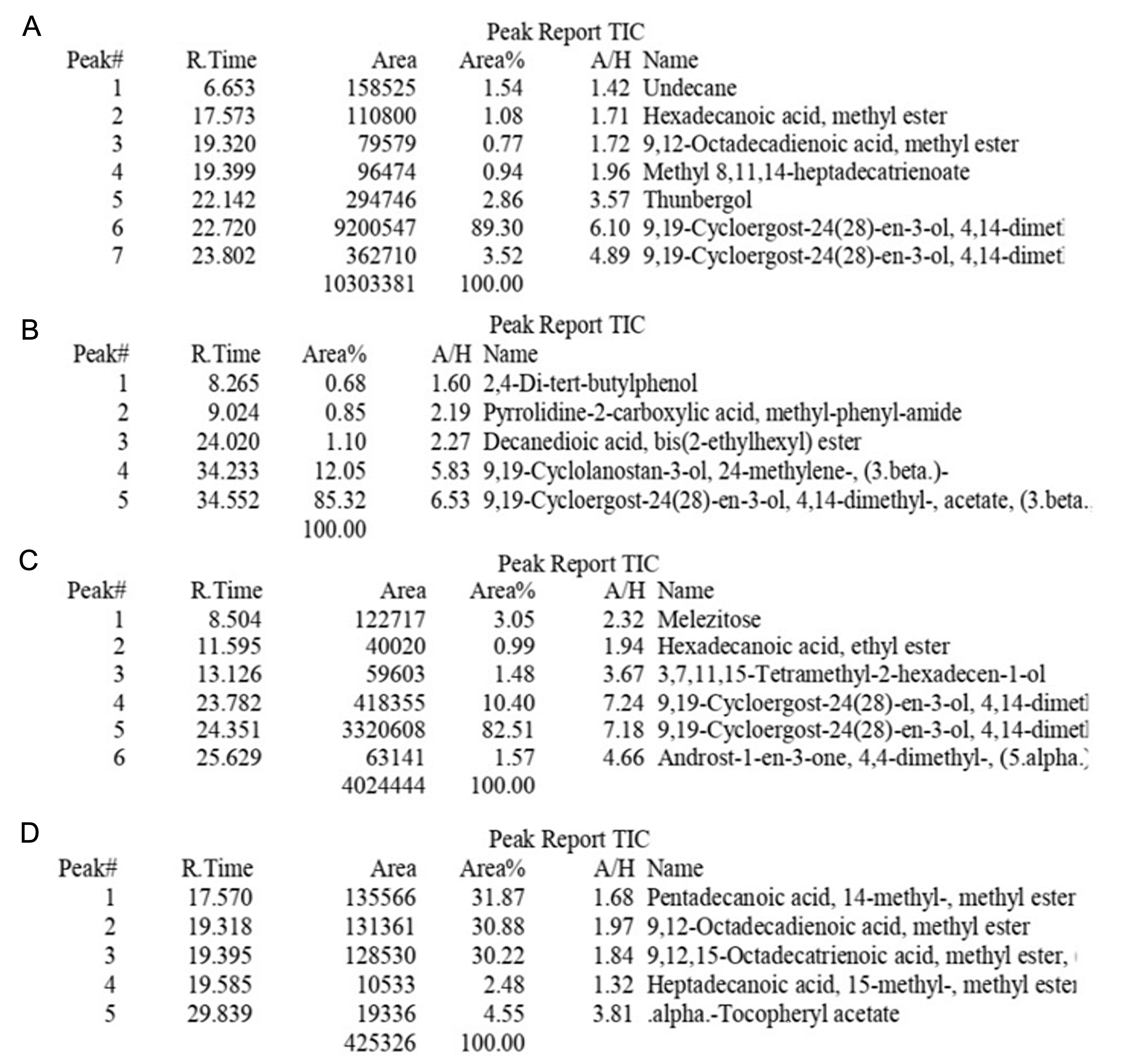{kind=link}
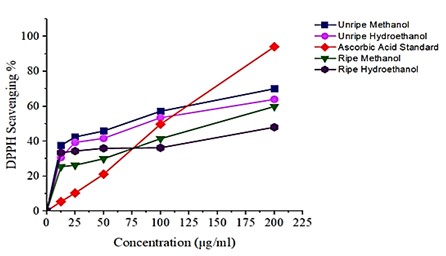{kind=link}
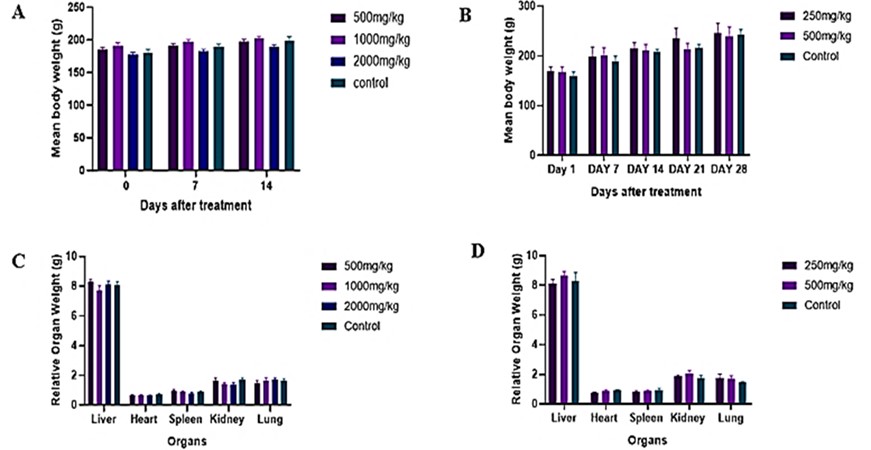{kind=link}
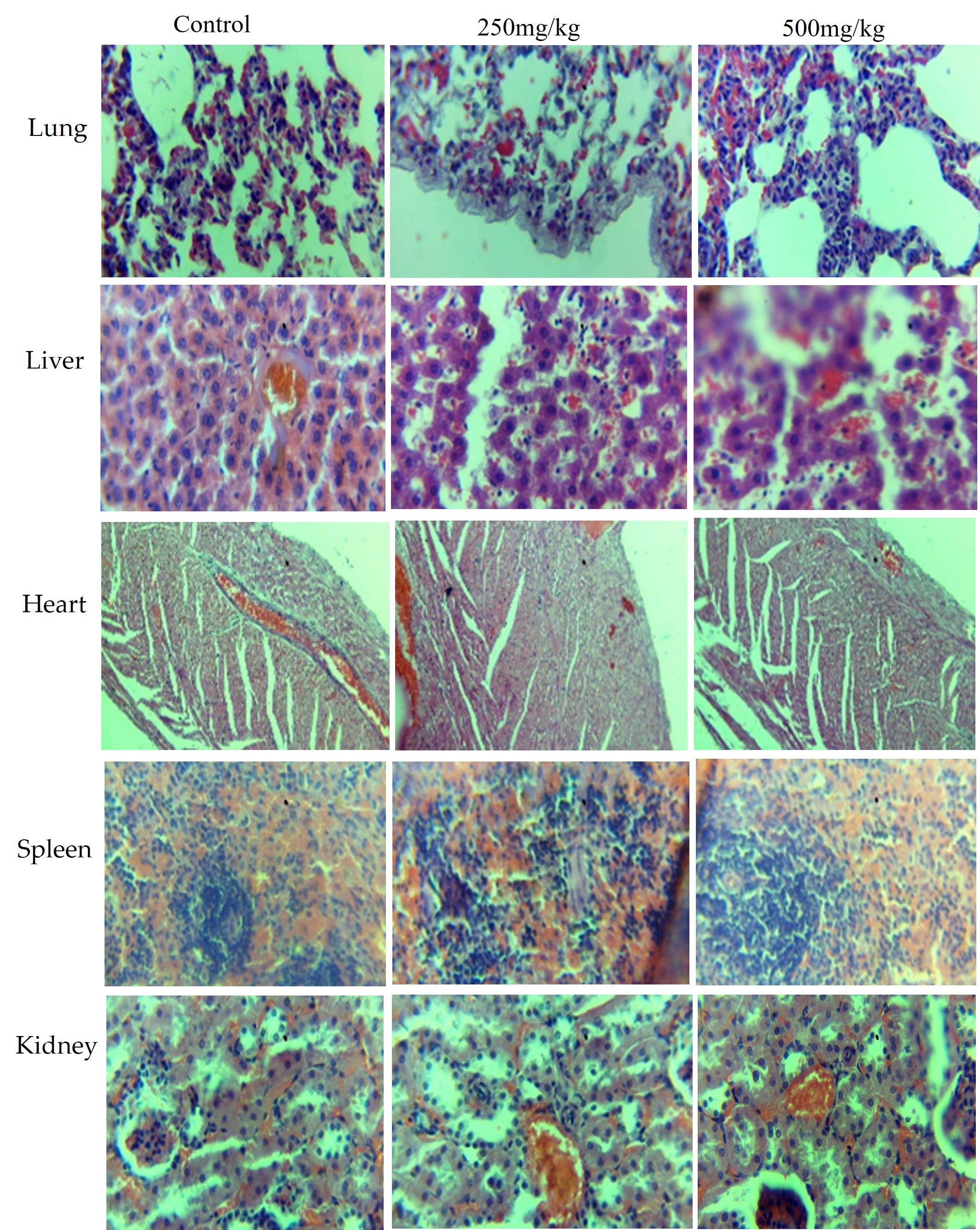{kind=link}